ЗАГАЛЬНА МІКРОБІОЛОГІЯ - Т.П. Пирог - 2004
7. СИСТЕМАТИКА ПРОКАРІОТ
7.5. ХАРАКТЕРИСТИКА ТАКСОНІВ ЗА ДЕВ’ЯТИМ ВИДАННЯМ КЕРІВНИЦТВА БЕРГІ З СИСТЕМАТИКИ БАКТЕРІЙ
7.5.4. Відділ Mendosicutes
Архебактерії — це переважно ґрунтові або водні мікроорганізми, які зустрічаються в анаеробних умовах у гіперсолоних, гідро- та геотермальних середовищах, а також як симбіонти у травному тракті тварин. До цієї групи входять аероби, анаероби, факультативні анаероби, здатні рости як хемолітоавтотрофи, гетеротрофи або факультативні гетеротрофи. Архебактерії можуть бути мезо- чи термофілами, причому деякі види здатні рости при температурі вище 100 С.
Унікальна біохімічна особливість архебактерій полягає в тому, що до складу ліпідів у них входять ізопренильні ефіри гліцерину. Відсутність муреїну (пептидоглікану, який вміщує мурамову кислоту) в клітинних стінках робить архебактерії стійкими до β-лактамних антибіотиків (пеніцилінів). Нуклеотидні послідовності 5S-, 16S- та 23S рРНК сильно відрізняються від відповідних у еубактерій та еукаріот.
Асиміляція вуглецю в автотрофних архебактерій відбувається не в циклі Кельвіна. СО2 фіксується через ацетил-КоА-шлях або через відновлювальний цикл трикарбонових кислот. Деякі архебактерії здатні фіксувати молекулярний азот. Серед коферментів та простетичних груп зустрічаються компоненти, які хоч і подібні до таких у еубактерій та еукаріот, але не ідентичним: похідне 5-деазарибофлавіну F420, нікельтетрапірольний фактор «мзо» метаноптерин, кофермент М.
Результати фарбування, за Грамом, можуть бути позитивними або негативними всередині одного порядку, оскільки типи клітинних оболонок дуже різняться. У грампозитивних видів клітинні стінки складаються з псеадомуреїну, метанохондрої- тинута гетерополісахариду. Грамнегативні клітини мають поверхневі шари, які складаються з глікопротеїну.
Форма клітин різноманітна: сферична, спіральна, пластинчаста, паличкоподібне. Зустрічаються одно- та багатоклітинні форми у вигляді ниток або агрегатів. Діаметр окремих клітин варіює від 0,1 до 15 мкм, а довжина ниток може досягати 200 мкм. Розмноження відбувається бінарним поділом, брунькуванням, перетяжкою, фрагментацією або невідомими способами. Забарвлення клітинної маси може бути червоним, пурпуровим, рожевим, оранжево-коричневим, жовтим, зеленим, темно- зеленим, сірим та білим.
Згідно з Керівництвом Бергі з ідентифікації бактерій (1997 р.) архебактерії розміщені в групах № 31-35 і поділяються на п'ять основних груп.
Метаногени (група 31). До них належать представники 18 родів (Methanobacterium, Methanococcus, Methanosarcina, Methanospirillum, Methanobrevibacterium, Methanogenium, Methanothrix та ін.). Слід зазначити, що у 1984 р. було описано лише 13 родів цих бактерій.
Строгі анаероби, здатні утворювати метан як основний кінцевий продукт метаболізму. Субстратами можуть бути Н2 + СО2, форміат, ацетат, метанол, метиламіни або Н2 + метанол. Сірку (S0) можуть відновлювати до H2S без одержання енергії. Мезо- L термофіли, нейтрофіли (оптимальне значення pH 7,0), зустрічаються галофільні види. Можуть давати синьо-зелену флуоресценцію при опроміненні світлом з довжиною хвилі 420 нм. Містять кофермент М, фактор 420, 430 і метаноптерин.
Сульфатредукуючі археї (група 32). До них належить рід Archaeoglobus, описано два види: Archaeoglobus fulgidus та Archaeo globus profundus. Дані про види Archaeoglobus відсутні в Bergey's Manual of Systematic Bacteriology. Вони описані у 1988-1990 рр.
Клітини кокоподібні неправильної форми, часто трикутні, розміщені поодиноко чи парами. Фарбуються, за Грамом, негативно. Утворюють зеленувато-чорні колонії діаметром 1-2 мм. Строгі анаероби, здатні утворювати H2S із сульфату в процесі дисиміляційної сульфатредукції. Крім того, утворюють невеликі кількості метану. Екстремальні термофіли (ріст при температурі до 92 С), діапазон pH 4,5-7,5 (оптимум близько 6), діапазон солоності — 0,9-3,6 % хлористого натрію. Здатні до хемолітотрофного, хемоорганотрофного або хемоміксотрофного росту. Для автотрофного росту необхідні тіосульфат і Н2. У гетеротрофних умовах використовують форміат, лактат, глюкозу, крохмаль і білки як донори електронів. В ультрафіолеті (довжина хвилі 420 нм) виявляють синьо-зелену флуоресценцію. Містять фактор 420 і метаноптерин, але не кофермент М та фактор 430.
Екстремально галофільні аеробні архебактерії (галобактерії) (група 33). Належать представники шести родів (Halococcus, Halobacterium, Haloarcula. Haloferox, Natronobacterium, Natronococcus). У 1984 p. було описано тільки три роди галобактерій — Halococcus, Haloarcula та Halobacterium.
Грамнегативні чи грампозитивні, аероби чи факультативні анаероби, хемоорганотрофи. Клітини паличкоподібні, мають форму від правильної до дуже неправильної. Потребують високої концентрації хлориду натрію (1,5 М і вище). Нейтрофіли чи алкалофіли. Мезофіли чи до деякої міри термофіли (до 55º С). Деякі види містять фотоактивний червоно-пурпуровий пігмент бактеріородо псин і здатні використовувати світло для синтезу АТФ.
Архебактерїї, які не мають клітинної стінки (група 34). Термоацидофіли, аероби (належить один рід — Thermoplasma). Оптимальна температура близько 60º С, оптимальне значення pH 1-2. Клітини кокоподібні, не мають клітинної стінки. Їх можна назвати термоацидофільними «мікоплазмами». Цитоплазматична мембрана містить багатий на манозу глікопротеїн і ліпоглікан.
Екстремальні термофіли і гіпертермофіли, які метаболізують S0 (група 35). Належать 14 родів (Acidianus, Desulfurolobus, SulfolobuSj Pyrobaculum, Pyrococcus, Desuifurococcus та ін.). У 1984 p. було описано лише сім родів.
Облігатні термофіли, аероби, факультативні анаероби чи строгі анаероби. Грамнегативні палички, нитки чи коки. Оптимальною для росту є температура в інтервалі 70-105 С. Ацидофіли та нейтрофіли. Автотрофи чи гетеротрофи. Більшість видів метаболізує сірку.
Деякі основні ознаки, які відрізняють еубактерії від архебактерії, наведені у табл. 7.3.
Таблиця 7.3
Деякі ознаки, що відрізняють еубактерії від архебактерій
Ознака |
Еубактерії |
Архебактерії |
|
Основні морфологічні та метаболічні ознаки |
|||
Строп анаероби, які утворюють метан (як основний кінцевий продукт) з H2 + СО2, форміату, ацетату, метанолу, метиламіну чи Н2 + метанолу. Виявляють синьо-зелену флуоресценцію при опроміненні світлом з довжиною хвилі 420 нм |
Ні |
Так |
|
Строгі анаероби, які утворюють Н2S з сульфату в процесі дисиміляційної сульфатредукції. Екстремальні термофіли (ріст при температурі до 92º С). Виявляють синьо-зелену флуоресценцію при опроміненні світлом з довжиною хвилі 420 нм |
Ні |
Так |
|
Грамнегативні чн грампозитивні, аероби чи факультативні анаероби, хемоорганотрофи. Клітини паличкоподібні, мають форму від правильної до дуже неправильної. Потребують високої концентрації хлориду натрію (1.5 М і вище). Нейтрофіли чн алкалофіли. Мезофіли чи до деякої міри термофіли (до 55 ºС). Деякі види містять фотоактивний червоно-пурпуровий пігмент бактеріородопсин і здатні використовувати світло для синтезу АТФ |
Ні |
Так |
|
Термоацидофіли, аероби, клітини кокоподібні, не мають клітинної стінки |
Ні |
Так |
|
Облігатні термофіли, аероби, факультативні анаероби чи строгі анаероби. Грамнегативні палички, нитчасті чи коки. Оптимальною для росту є температура в інтервалі 70-105 º С. Ацидофілії та нейтрофіли. Автотрофи чи гетеротрофи. Більшість видів метаболізує сірку |
Ні |
Так |
|
Клітинна стінка (якщо є) |
|||
Містить мурамову кислоту |
Так |
Ні |
|
Чутливість до антибіотиків |
|||
Чутливі до пеніциліну або його аналогів, які пригнічують синтез пептидоглікану, що містить мурамову кислоту |
Так |
Ні |
|
Ліпіди |
|||
У складі фосфоліпідів мембран виявлено: довголанцюгові спирти (фітаноли), зв’язані ефірними зв’язками з гліцерином з утворенням фітанільних (Сж) диефірів гліцерину або дифітанільних (С10) дигліцеринових тетреефірів |
Ні |
Так |
|
Шлях біосинтезу ліпідів: мевалонатний |
Ні |
Так |
|
малонатний |
Так |
Ні |
|
Молекулярно-біологічні ознаки |
|||
До складу нуклеотидів ТТС-петлі молекул тРНК входить; риботимідин |
Так |
Ні |
|
псевдоурідин та 1-метилпсевдоурідин |
Ні |
Так |
|
Амінокислота, з якої починається поліпептидний ланцюг під час синтезу білка: метионін |
Ні |
Так |
|
N-формілметионін |
Так |
Ні |
|
Аміноацильна гілка ініціаторноі тРНК закінчується парою основ «АУ» |
Ні |
Так |
|
Синтез білка на рибосомах пригнічується: |
|||
анізоміцином |
Ні |
Так |
|
канаміцином |
Так |
Ні |
|
хлорамфеніколом |
Так |
Ні |
|
Фактор елонгації EF-2 АДФ рибозилюється дифтерійним токсином |
Ні |
Так |
|
Фактор елонгації EF-2 містить амінокислоту дифтамід |
Ні |
Так |
|
Деякі гени тРНК містять інтронн |
Ні |
Так |
|
ДНК-залежні РНК-полімерази: |
|||
являють собою мультиферментний комплекс |
Ні |
Так |
|
пригнічуються рифампіцином і стрептолідигіном |
Так |
Ні |
|
Відкриття архебактерій (1977 р.) спричинило дискусію про їх місце в системі живих істот і повернуло до проблеми клітинної еволюції. Нині існують два уявлення про шляхи клітинної еволюції (рис. 7.1).
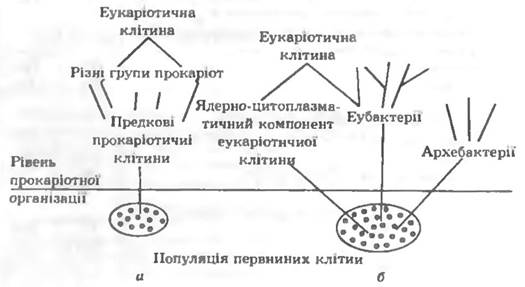
Рис. 7.1. Можливі шляхи клітинної еволюції: и монофілітичний: б — поліфілетичний
За одним з них (рис. 7.1, а) з популяції первинних клітин виникла популяція предкових прокаріотних клітин, з якої утворилися різні групи прокаріот (монофілетична група). Еукаріотична клітина виникла в результаті ендосимбіозу прокаріотичної клітини хазяїна та ендосимбіонту. Такими клітинами-хазяями могли бути архебактерії, що утворилися з предкових прокаріотних клітин на ранньому етапі їх існування і поглинули вільно існуючі прокаріоти, які потім стали хлоропластами та мітохондріями еукаріот. За іншим уявленням (рис. 7.1, б)еубактеріі, архебактерії і ядерно-цитоплазматичний компонент еукаріотичної клітини (мітохондрії, хлоропласти) дивергували від спільного предка самостійно, незалежно і досягли рівня організації сучасних прокаріотних клітин (поліфілетичне походження).