Общая микробиология - Шлегель Г. 1987
Грибы
Фикомицеты (низшие грибы)
К низшим грибам мы относим большую группу грибов, вегетативные тела которых (даже при сильном ветвлении гиф) не имеют перегородок и потому многоядерны. Такой таллом называют ценоцитным. У большинства низших грибов споры образуются в спорангиях. Примитивные формы, приспособившиеся к жизни в воде, образуют подвижные споры и гаметы. При переходе от водных форм к более совершенным земноводным и наземным подвижные стадии встречаются только изредка.
Хитридиомицеты. Это главным образом водные грибы, хотя некоторые представители хитридиомицетов встречаются и в почве. Все они микроскопически малы. Клеточная стенка у них состоит, по-видимому, в основном из хитина. Многие хитридиомицеты паразитируют на планктонных водорослях и высших водных растениях. Как экономически важного паразита культурных растений следует отметить Synchytriun endobioticum - возбудителя рака картофеля. Rhizophidium pollinis - xитридиомицет, паразитирующий на пыльце сосны, - является излюбленным демонстрационным объектом.
Оомицеты. Эта группа включает водные и наземные формы, размножающиеся бесполым путем-при помощи зооспор, снабженных двумя жгутиками. Saprolegnia и Leptomitus, известные под общим названием «водяная плесень», живут в воде. Peronosporales перешли к наземному образу жизни. Это облигатные паразиты, весь цикл их развития проходит в организме высших растений; однако они еще образуют зооспоры. К оомицетам принадлежат некоторые из самых вредоносных патогенов, вызывающих опустошительные эпифитотии, в частности Phytophthora infestans (возбудитель фитофтороза картофеля) и Plasmopara viticola (возбудитель ложной мучнистой росы винограда).
Широко распространена Saprolegnia, легко поддающаяся выделению и культивированию. Методом «приманки» нетрудно получить ее накопительную культуру, если в чашку, заполненную прудовой водой, положить мертвую муху таким образом, чтобы она лежала с распростертыми крыльями (ногами вниз) на поверхности воды. Уже через несколько дней все тельце мухи будет покрыто гифами и спорангиями. Под микроскопом хорошо видно, как образуются спорангии и как выскальзывают из них зооспоры (рис. 5.4).
У Saprolegnia и родственных видов интересно такое явление, как двустадийность образования зооспор - диплания. Вышедшие из спорангия первичные зооспоры, поплавав некоторое время, останавливаются и превращаются в цисты; из цисты выходит новая зооспора, которая вскоре тоже инцистируется; и только эта циста прорастает, образуя ростовую трубку и гифы.
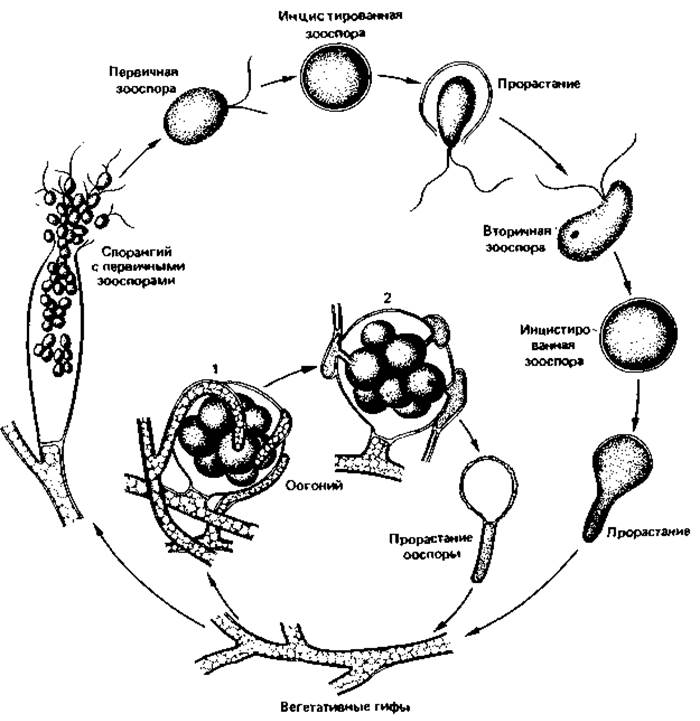
Рис. 5.4. Цикл развития Saprolegnia sp. 1 - оогоний с шаровидными оосферами и антеридии; 2 - то же в поперечном разрезе; видны оплодотворяющие трубки антеридиев, внедрившиеся в оогоний. (Alexopoulos С. J., Einführung in die Mykologie, Stuttgart: Fischer, 1966.)
Половое размножение осуществляется при непосредственном контакте гаметангиев, т. е. антеридия и оогония. Большинство Saprolegniaceae гомоталличны: оогонии и оплодотворяющие их антеридии образуются на одном и том же вегетативном теле. Оогонии имеют шаровидную форму, толстые стенки и содержат несколько яйцеклеток (оосфер). Антеридии меньше по своим размерам и образуются на кончиках гиф. Зрелые антеридии (по одному или по нескольку) прикрепляются к оогонию. Их оплодотворяющие трубки проникают к оосферам через стенку оогония. Когда одно из ядер антеридия достигнет соответствующего ядра яйцеклетки и сольется с ним, образуется диплоидное ядро зиготы. Оплодотворенная оосфера окружается толстой стенкой и превращается в ооспору. После длительного периода покоя ооспоры прорастают; при этом происходит редукционное деление. Цикл завершается образованием нового спорангия.
Зигомицеты. Название «зигомицеты» (от греч. «зигос» - ярмо) связано с особым типом полового размножения, при котором образуются зигоспоры. Ценозигота, или зигоспора, образуется в результате слияния двух гаметангиев (гаметангиогамия), которые в виде мостика («ярма») соединяют две родительские гифы (рис. 5.6). Зигомицеты - наиболее высокоразвитая, перешедшая к наземному образу жизни группа фикомицетов. Их подразделяют на три порядка: Mucorales, Entomophthorales и Zoopagales. Мы здесь рассмотрим только первый из них.
Представители порядка Mucorales (мукоровые, головчатые плесени) живут на гниющих органических материалах; некоторые из них - копрофилы, т. е. предпочтительным субстратом служит для них навоз. Именно поэтому лошадиный навоз и навозная жижа сыграли в свое время немаловажную роль, послужив субстратом при проведении очень ценных исследований. Такие грибы, как Mucor mucedo (обычная головчатая плесень), Rhizopus nigricans (вульгарная хлебная плесень), R. oryzae, R. arrhizus, R. rouxii, Phycomyces blakesleeanus, Choanephora cucurbitarum, Blakesleea и ряд других, известны отнюдь не только микологам. В связи с той ролью, которую играют эти грибы в промышленности, ими интересуются также химики и биотехнологи.
В анаэробных условиях грибы растут, как правило, очень плохо и лишь короткое время. В отсутствие молекулярного кислорода они переходят к брожению; многие из них образуют в таких случаях молочную кислоту или этиловый спирт. Изменяется при этом и форма роста: Mucor racemosus, например, образует в анаэробных условиях почкующийся мицелий, и его молодые клетки, подобно дрожжам, размножаются почкованием.
Распространение мукоровых грибов происходит очень быстро как с помощью многочисленных спорангиоспор, так и благодаря быстрому росту гиф. Rhizopus stołonifer (= R. nigricans) образует, например, «усы», или столоны, тянущиеся на довольно значительные расстояния (измеряемые сантиметрами).
Для бесполого размножения служат спорангии и спорангиоспоры. Из обильно растущего мицелия поднимаются вертикально боковые ветви - спорангиеносцы. Кончики их отделяются поперечной перегородкой и вздуваются. Происходит расслоение на две зоны - богатую содержимым наружную и бедную содержимым внутреннюю, которые затем разделяются клеточной стенкой. Поперечная перегородка при этом куполообразно изгибается кверху, образуя так называемую колонку (columella) - как бы продолжение спорангиеносца внутри спорангия. В периферической зоне спорангия могут находиться сотни или тысячи ядер, и вокруг каждого из них скапливается некоторое количество цитоплазмы - образуются спорангиоспоры. У некоторых представителей мукорових спорангиеносцы разветвленные, а спорангии мелкие. В таких спорангиях (спорангиолах) содержится по нескольку спор или всего одна спора.
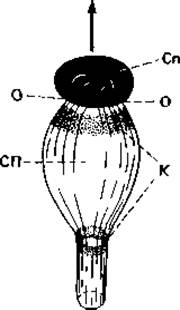
Рис. 5.5. Спорангиеносец фикомицета Pilobolus. Стрелкой показано направление, в котором отбрасывается спорангий (Сn) после отрыва от субспорангиального пузыря (СП); О - место отрыва; К - зона цитоплазмы, богатая каротиноидами. (Nultsch W., Allgemeine Botanik, 3 Aufl., Stuttgart: Thieme, 1968.)
У Pilobolus спорангий имеет несколько иную форму (рис. 5.5). У этого гриба несущая гифа под спорангием вздувается, образуя пузырь. На этом вздутии, как круглая крышка, сидит спорангий. За счет впитывания большого количества воды внутреннее давление резко возрастает, так что при созревании спорангиеносец в верхней вздутой части разрывается и спорангий (вместе с колонкой) отбрасывается на высоту до 2 м (отсюда и название Pilobolus, что означает «метатель снаряда»). При этом направление полета спорангия не случайно: спорангиеносец обладает положительным фототропизмом, т. е. растет в сторону источника света, а потому и спорангий летит в том же направлении. Pilobolus образует спорангии только на среде, содержащей вытяжку из навоза. Необходимым фактором роста для него является копроген - вещество из группы сидераминов.
Половое размножение и цикл развития мукоровых грибов мы рассмотрим на примере Rhizopus nigricans (рис. 5.6). Спорангиоспоры у этого вида многоядерные. При благоприятных условиях они прорастают, образуя сильно разветвленный воздушный мицелий. Там, где гифы соприкасаются с субстратом, формируются ризоиды, которые проникают в субстрат. Непосредственно над этим местом появляются один или несколько спорангиеносцев. У R. nigricans половое размножение возможно лишь в тех случаях, когда встречаются два физиологически различных совместимых мицелия, т. е. (+)-штамм и (-)-штамм. При сближении образуются копуляционные ветви, которые, вздуваясь, превращаются в прогаметангии, обогащаются цитоплазмой (число ядер при этом тоже увеличивается) и в конце концов отделяются от несущих гиф поперечными перегородками. В зоне контакта гаметангиев происходит лизис клеточных стенок, и протопласты сливаются (гаметангиогамия). За этим следует попарное соединение (+)- и (-)-ядер, а потом их слияние. Ценозигота увеличивается в размерах и в конце концов превращается в толстостенную зигоспору. После некоторого периода покоя зигоспора прорастает с образованием зародышевого спорангия; ядра при этом претерпевают редукционное деление. Таким образом, вегетативное тело гриба представляет собой гаплоидную фазу.
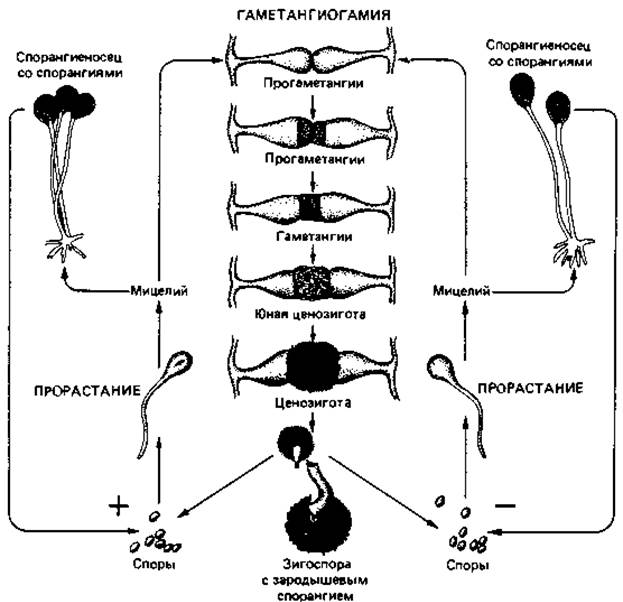
Рис. 5.6. Цикл развития Rhizopus nigricans.
Некоторые представители мукоровых гомоталличны и способны к самооплодотворению.