Общая микробиология - Шлегель Г. 1987
Основные механизмы обмена веществ и преобразования энергии
Пути катаболизма гексоз
Несколько путей ведут от глюкозы к С3-соединениям и среди них к пирувату - одному из важнейших промежуточных продуктов метаболизма. Чаще других используется путь распада через образование фруктозо-1,6-бисфосфата; его называют фруктозобисфосфатным путем, гликолитическим расщеплением, гликолизом или (по имени изучивших его исследователей) путем Эмбдена-Мейергофа-Парнаса (рис. 7.3). Другой ряд реакций, к осуществлению которых способно большинство организмов, образует цикл, известный под названием окислительного пентозофосфатного пути, уексозомонофосфатного пути или схемы Варбурга-Диккенда-Хореккера (рис. 7.4). Обратная последовательность; реакций этого пути включает важные этапы, ведущие к регенерации акцептора СO2 при автотрофной фиксации углекислоты. Только у бактерий встречается, видимо, путь Энтнера - Дудорова, или, как его еще называют, КДФГ-путь (по характерному промежуточному продукту- 2-кето-3-дезокси-6-фосфоглюконату, КДФГ; рис. 7.5). Другие сходные механизмы распада гексоз имеют более специальное значение.
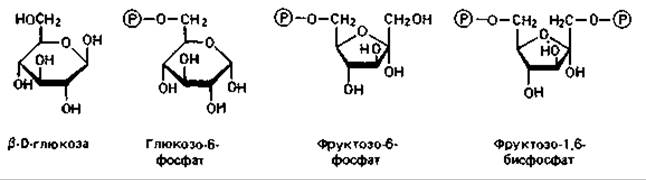
Глюкоза в клетке сначала фосфорилируется обычно в положении 6 при участии гексокиназы как катализатора и АТР как донора фосфата. Глюкозо-6-фосфат представляет собой метаболически активную форму глюкозы в клетке и служит исходным пунктом для любого из трех упомянутых путей распада.
7.2.1 Фруктозо-1,6-бисфосфатный путь (гликолиз)
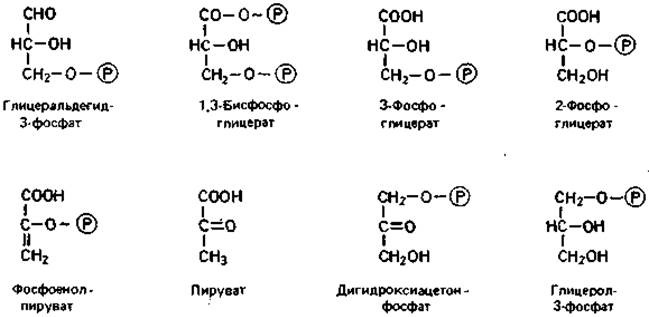
На фруктозобисфосфатном пути (рис. 7.3) глюкозо-6-фосфат при подготовке к расщеплению изомеризуется глюкозофосфат-изомеразой в фруктозо-6-фосфат; затем происходит фосфорилирование в положении 1 под действием фосфофруктокиназы за счет АТР. Образовавшийся фруктозо- 1,6-бисфосфат расщепляется фруктозобисфосфат-алъдолазой до дигидроксиацетонфосфата и глицеральдегид-3-фосфата. Оба триозофосфата находятся в равновесии между собой; установление этого равновесия катализируется триозофосфат-изомеразой. Дигидроксиацетонфосфат может восстанавливаться глицеролфосфат-дегидрогеназой до глицерол-фосфата, который гидролизуется глицерол-1-фосфатазой с образованием глицерола и ортофосфата. Обычно же сначала происходит превращение образовавшегося под действием альдолазы дигидроксиацетон- фосфата в глицеральдегид-3-фосфат, который затем окисляется.
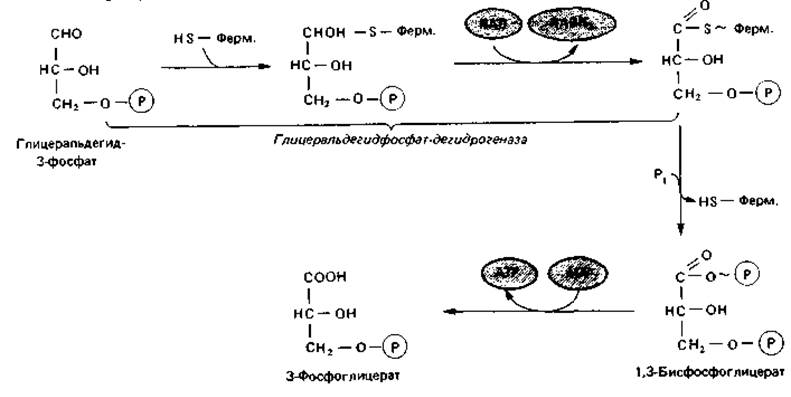
Последующее дегидрирование представляет собой с энергетической стороны важнейший этап данного пути, а также других путей, приводящих к образованию глицеральдегид-3-фосфата. Часть энергии, освобождающейся при окислении глицеральдегид-3-фосфата в 3-фосфоглицерат (AG0' = —67 кДж), сохраняется в форме высокоэнергетического фосфата. Сначала происходит присоединение альдегидной группы к SH- группе глицеральдегидфосфат-дегидрогеназы, а затем отщепление водорода, который переносится на NAD. Образовавшийся ацил-8-фермент представляет собой тиоэфир, богатый энергией. В результате фосфоролиза (при котором ацильная группа отделяется от фермента с присоединившимся к ней ортофосфатом) эта энергия сохраняется в 1,3-бисфосфо-глицерате. При участии фосфоглицераткиназы богатая энергией фосфатная группа переносится на ADP с образованием 3-фосфоглицерата и АТР. Такого рода процесс называют фосфорилированием на уровне субстрата. Для предшествующего окисления глицеральдегид-3-фосфата наряду с ферментом необходимы также ортофосфат и ADP. В случае их отсутствия расщепление глюкозы на этом уровне прекращается. Это имеет значение для регуляции процесса распада глюкозы («эффект Пастера»).
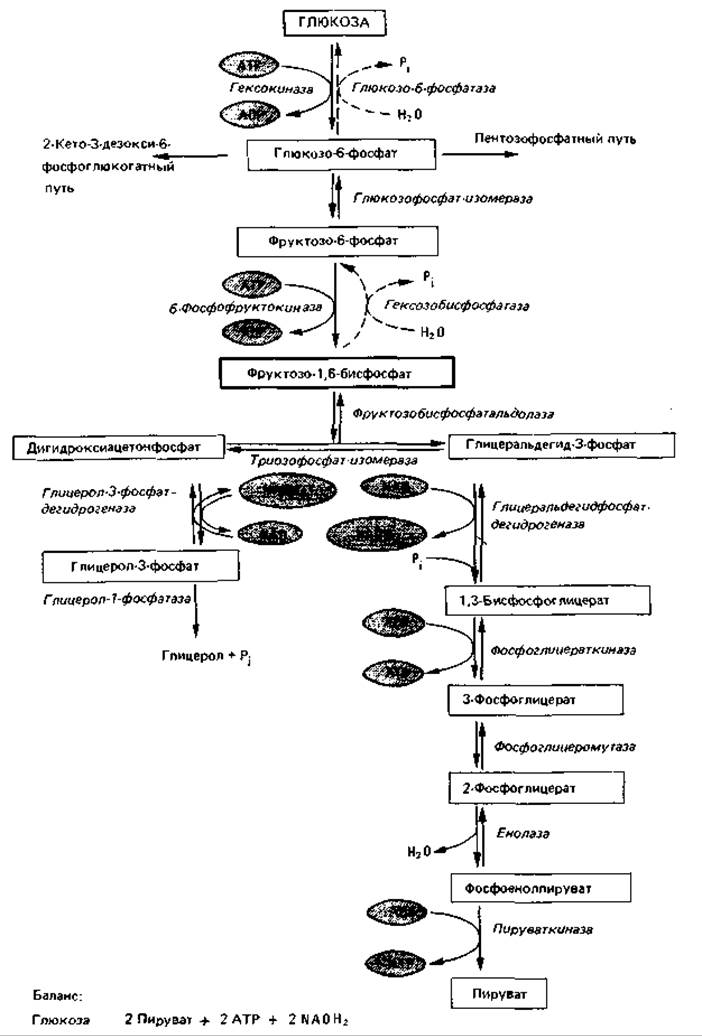
Рис. 7.3. Фруктозо-1,6-бисфосфатный путь расщепления глюкозы (гликолиз).
Под действием фосфоглицеромутазы 3-фосфоглицерат превращается в 2-фосфоглицерат, из которого в результате отнятия воды (катализируемого енолазой) образуется фосфоенолпируват. Это тоже высокоэнергетический фосфат, с которого богатая энергией фосфатная группа переносится пируваткиназой на ADP и таким образом сохраняется. Образующийся при этом пируват служит исходным пунктом дальнейших процессов расщепления, преобразования и синтеза. Все реакции фруктозо-1,6-бисфосфатного пути, за исключением трех (гексокиназной, 6-фосфофруктокиназной и пируваткиназной), полностью обратимы.
Если весь тризофосфат, образовавшийся в результате расщепления фруктозо-1,6-бисфосфатного пути, за исключением трех (гексокиназной, катаболизма глюкозы по фруктозо-1,6-бисфосфатному пути слагается из двух молекул пирувата, двух (4 минус 2) молекул АТР и двух молекул NADH.
Обе реакции, протекающие с выделением энергии при превращении триозофосфата в пируват, служат для анаэробных организмов важнейшими этапами, доставляющими энергию. В анаэробных условиях все микроорганизмы, сбраживающие углеводы (за немногими исключениями), используют энергию, получаемую в результате окисления глицеральдегидфосфата в пируват.
7.2.2 Пентозофосфатный путь
В пентозофосфатном пути (рис. 7.4) глюкозо-6-фосфат дегидрируется глюкозо-6-фосфат-дегидрогеназой; при этом водород переносится на NADP и образуется 6-фосфоглюконолактон, который спонтанно или при участии фермента (глюконолактоназы) гидролизуется до 6-фосфоглюконата. Этот последний дегидрируется дегидрогеназой до 3-кето-6-фосфоглюконата, из которого затем путем декарбоксилирования образуется рибулозо-5-фосфат. Этим завершается собственно процесс окисления.
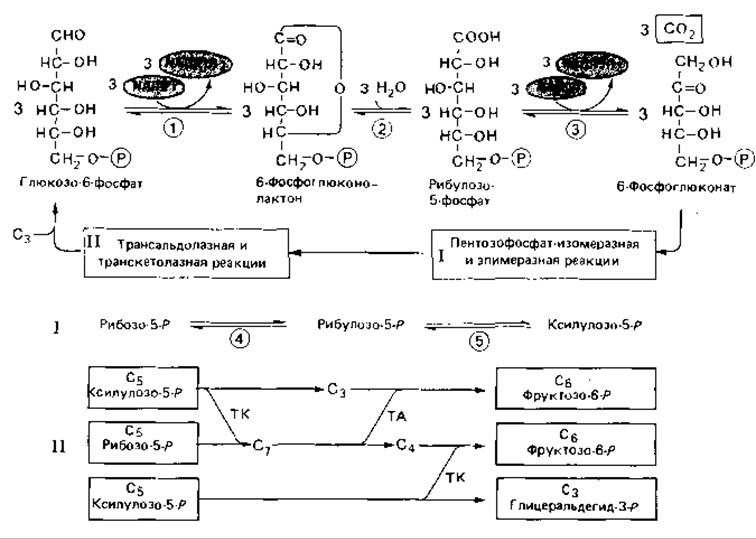
Рис. 7.4. Пентозофосфатный путь окислительного расщепления глюкозо-6-фосфата. Вверху представлен пентозофосфатный цикл. Окислительные этапы заканчиваются образованием рибулозо-5-фосфата. Рибу лозо-5-фосфат находится в равновесии с рибозо-5-фосфатом и ксилулозо-5-фосфатом, что обеспечивается ферментативным катализом. Пентозофосфаты при участии транскетолазы и трансальдолазы превращаются в два фруктозофосфата и один глицеральде-гидфосфат. Эта последовательность реакций полностью обратима и в обратном направлении включается в рибулозобисфосфатный цикл фиксации СO2, в рибулозомонофосфатный цикл фиксации формальдегида и другие циклы. Участвующие ферменты (цифры в кружках): 1 - глюкозо-6-фосфатдегидрогеназа; 2 - лактоназа; 3 - 6-фосфоглюконатдегидрогеназа; 4 - фосфорибозоизомераза; 5 - рибулозо-5-фосфат-3-эпимераза; 6 - транскетолаза; 7 - трансальдолаза.
Последующие реакции надо рассматривать только как процессы превращения пентозофосфатов в гексозофосфаты и обратно. Благодаря включению такой последовательности реакций окислительный пентозофосфатный путь замыкается в цикл. Рибулозо-5-фосфат находится в равновесии с рибозо-5-фосфатом и ксилулозо-5-фосфатом. Рибозофосфат - важный предшественник в процессе синтеза нуклеотидов и нуклеиновых кислот. При участии транскетолазы и трансальдолазы пентозофосфаты превращаются в две молекулы фруктозо-6-фосфата и одну молекулу глицеральдегид-3-фосфата. В результате изомеризации фруктозо-6-фосфата в глюкозо-6-фосфат и конденсации двух молекул триозофосфата в гексозофосфат все перечисленные реакции замыкаются в один цикл, при одном обороте которого из трех молекул глюкозо-6-фосфата образуются две молекулы фруктозо-6-фосфата, одна молекула глицеральдегид-3-фосфата, три молекулы СО2 и трижды по две молекулы NADPH2. Ферменты глюкозо-6-фосфат-дегидрогеназа и фосфоглюконатдегидрогеназа у многих (если не у большинства) бактерий переносят водород с субстратов не только на NADP, но и на NAD.
Описанный цикл представляет собой явно побочный путь, значение которого следует видеть в подготовке важных исходных веществ (пентозофосфатов, эритрозофосфата, глицеральдегид-3-фосфата), а также получении восстановительных эквивалентов (NADPH2) для процессов синтеза. Пентозофосфаты - предшественники нуклеотидов и нуклеиновых кислот-образуются путем дегидрирования и декарбоксилирования глюкозо-6-фосфата, а также в транскетолазной и трансальдолазной реакциях из фруктозо-6-фосфата.
7.2.3 2-Кето-3-дезонси-6-фосфоглюнонатный путь
Глюкозо-6-фосфат сначала, как это было описано выше для пентозофосфатного пути, дегидрируется до 6-фосфоглюконата. Под действием фосфоглюконатдегидратазы от него отщепляется вода и образуется 2-кето-3-дезокси-6-фосфоглюконат (рис. 7.5). Кетодезоксифосфоглюконат расщепляется специфической альдолазой на пируват и глицеральдегид-3-фосфат. Последний окисляется до пирувата, так же как и в фруктозобисфосфатном пути.
В отношении образования ATP, NADH2 и NADPH2 между описанными путями катаболизма сахаров имеются существенные различия. На каждый моль глюкозы, окисляемой до пирувата по фруктозобисфосфатному пути, образуется 2 моля АТР и 2 моля NADH2, а при 2-кето-З- дезокси-6-фосфоглюконатном пути - по одному молю ATP, NADH2 и NADPH2. Таким образом, в последнем случае 1 моль АТР и 1 моль NADH2 заменяются на один моль NADPH2, что эквивалентно. Эта эквивалентность согласуется с данными о том, что перенос водорода с NADH2 на NADP с участием трансгидрогеназы во многих случаях требует затраты энергии и протекает с использованием АТР.
Микроорганизмы заметно различаются между собой по степени использования того или иного из рассмотренных путей (табл. 7.3). Ферменты фруктозобисфосфатного пути, как правило, являются обязательными компонентами клетки, хотя у многих бактерий этот путь действует лишь в обратном направлении (необратимые этапы катализируются при этом другими ферментами). Пентозофосфатный путь тоже, по-видимому, имеет универсальное значение. 2-Кето-3-дезокси-6-фосфоглюконатный путь у бактерий очень широко распространен; он имеет принципиальное значение для использования глюконата. Например, в то время как глюкоза у Escherichia coli и видов Clostridium расщепляется по фруктозобисфосфатному пути, глюконат включается у них в промежуточный обмен через 2-кето-3-дезокси-6-фосфоглюконатный путь.
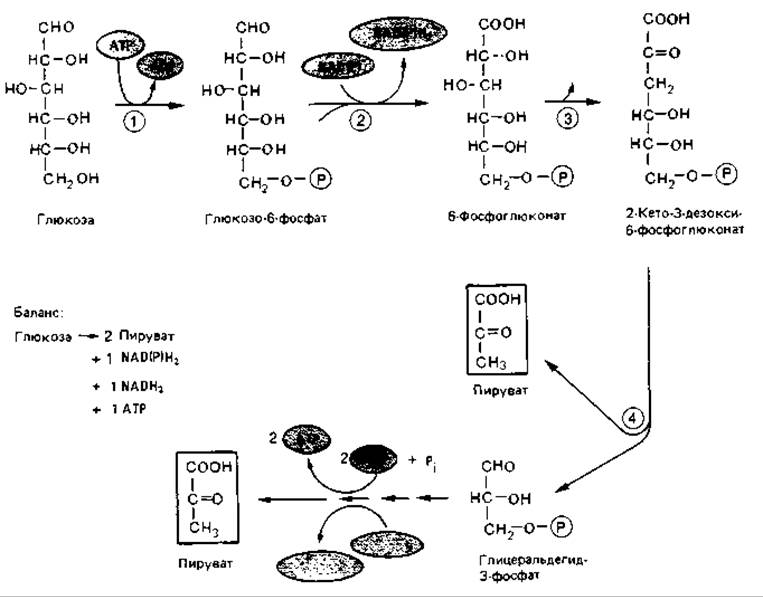
Рис. 7.5. 2-Кето-3-дезокси-6-фосфоглюконатный путь окислительного расщепления глюкозы (путь Энтнера-Дудорова). Участвующие ферменты (цифры в кружках): 1 - гексокиназа; 2 - глюкозо-6-фосфатдегидрогеназа; 3 - фосфоглюконатдегидрогеназа; 4 - фосфо-2-кето-3-дезоксиглюкопат-альдолаза. На схеме не показан 6 -фосфоглюконолактон, образующийся как промежуточный продукт при дегидрировании глюкозо-6-фосфата.
Таблица 7.3. Участие различных путей в катаболизме гексоз (в процентах)
|
Вид |
Фруктозо-1,6-бисфосфатный путь |
Пентозофосфатный путь |
2-Кето-3-дезокси-6-фосфоглюконатный путь |
|
Candida utilis |
70-80 |
30-20 |
|
|
Streptomyces griseus |
97 |
3 |
|
|
Penicillium chrysogenum |
77 |
23 |
|
|
Escherichia coli |
72 |
28 |
|
|
Bacillus subtilis |
74 |
26 |
|
|
Pseudomonas aeruginosa |
29 |
71 |
|
|
Gluconobacter oxydans |
100 |
||
|
Pseudomonas saccharophila |
100 |
||
|
Alcaligenes eutrophus |
100 |
Клостридии и некоторые аэробные бактерии используют для катаболизма глюконата особый вариант 2-кето-3-дезокси-6-фосфоглюконатного пути: сначала глюконат под действием глюконатдегидратазы превращается в 2-кето-3-дезоксиглюконат, и только на этом этапе происходит его фосфорилирование за счет АТР при участии кетодезоксиглюконокиназы; 2-кето-3-дезокси-6-фосфоглюконат расщепляется под действием фосфо-2-кето-3-дезоксиглюконатальдолазы.
7.2.4 Окисление пирувата
Пируват занимает центральное положение в промежуточном метаболизме и может служить предшественником разнообразных продуктов. Многие организмы окисляют большую часть образующегося в ходе катаболизма пирувата до ацетилкофермента А. Важнейшую роль у бактерий играют три реакции:

Реакция (1) катализируется мультиферментным комплексом пируват-дегидрогеназой. Этот фермент имеется почти у всех аэробных организмов и служит главным образом для образования ацетил-СоА, который поступает затем в цикл трикарбоновых кислот (рис. 7.6); его функция будет подробно описана ниже.
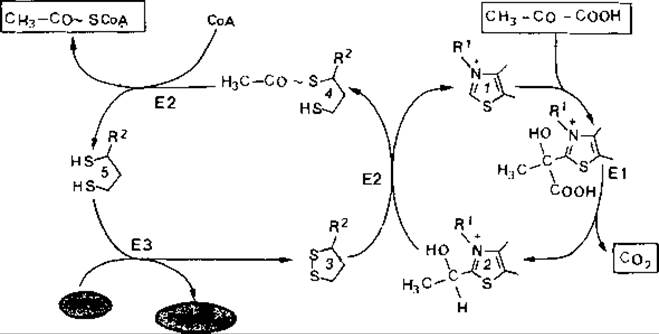
Рис. 7.6. Схема реакций, участвующих в дегидрировании пирувата (пояснения в тексте).
Реакцию (2) катализирует пируват: ферредоксин-оксидоредуктаза — фермент, который у многих анаэробных бактерий (например, у клостридиев) имеет особое значение.
Реакцию (3) катализирует пируват: формиат-лиаза. Этот фермент есть у многих анаэробных бактерий, выделяющих муравьиную кислоту («муравьинокислое брожение», разд. 8.4), в частности у Enterobacteriaceae, но встречается также у фототрофных бактерий.
У дрожжей и некоторых бактерий, образующих этанол, имеется четвертый фермент, окисляющий пируват:
(4) Пируват → Ацетальдегид + СО2
Этот фермент - пируватдекарбоксилазарасщепляет пируват на ацетальдегид и СО2. Ацетальдегид затем восстанавливается до этанола.
Дегидрирование пирувата пируватдегидрогеназой. Этот мультиферментный комплекс при участии кофакторов превращает пируват в ацетилкофермент А и СО2 [см. реакцию (1) выше и рис. 7.6].
В состав мультиферментного комплекса вхолят три белка: пируватдегидрогеназа (El), дигидролипоамид-трансацетилаза (Е2) и дигидролипоамид-дегидрогеназа (Е3):
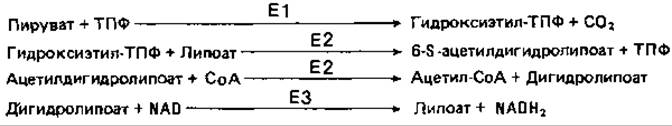
На начальном этапе (E1) пируват взаимодействует со 2-м углеродным атомом тиазольного кольца (1) тиаминпирофосфата (ТПФ), в результате чего отщепляется СО2. Образующийся гидроксиэтил-ТПФ (2) реагирует с присоединенным к Е2 липоатом (3), который при этом восстанавливается, а у вторичной SH-группы удерживает ацетильный остаток (4). Затем Е2 катализирует перенос ацетильной группы на кофермент А: при этом остается дигидролипоат (5), который снова окисляется при помощи Е3 до липоата при одновременном восстановлении NAD.
У строго анаэробных бактерий пируватдегидрогеназный мультиферментный комплекс отсутствует.