Общая микробиология - Шлегель Г. 1987
Перенос электронов в анаэробных условиях
Денитрификация и восстановление нитрата
В результате фосфорилирования, сопряженного с переносом электронов при дыхании аэробных организмов, образуется намного больше АТР, чем при фосфорилировании на уровне субстрата при брожении; поэтому неудивительно, что в процессе биохимической эволюции возник и в дальнейшем сохранился такой тип метаболизма, при котором водород от органического субстрата переносится на «связанный кислород». При этом «носителями» кислорода могут быть нитрат, сульфат, карбонат или другие соединения; они восстанавливаются водородом субстрата. Способность переносить электроны на эти соединения дает бактериям возможность окислять субстраты без участия молекулярного кислорода и таким образом извлекать больше энергии, чем это возможно при брожении.
Такого рода бактерии обладают системой переноса (транспорта) электронов и, как правило, содержат цитохромы. Получение энергии путем фосфорилирования, сопряженного с переносом электронов, при участии указанных выше «носителей кислорода» (вернее, конечных акцепторов водорода) в принципе сходно с дыханием, при котором роль конечного акцептора водорода играет кислород. Но поскольку такой процесс осуществляется в анаэробных условиях, говорят об анаэробном дыхании, причем различают нитратное, сульфатное, карбонатное дыхание и т.д. (рис. 9.1). Бактерии, способные к анаэробному дыханию с использованием неорганических акцепторов водорода (нитрат, сульфат, карбонат), играют очень важную роль как в природе, так и в хозяйстве человека.
Микроорганизмы используют нитрат для двух целей. Во-первых, подобно большинству растений, многие бактерии способны извлекать из него азот для синтеза азотсодержащих клеточных компонентов. Такая ассимиляционная нитратредукция может протекать и в аэробных, и в анаэробных условиях. Во-вторых, возможна также диссимиляционная нитратредукция, или «нитратное дыхание»; при этом нитрат в анаэробных условиях служит конечным акцептором водорода. В обоих случаях нитрат сначала восстанавливается до нитрита с помощью молибденсодержащего фермента нитратредуктазы.
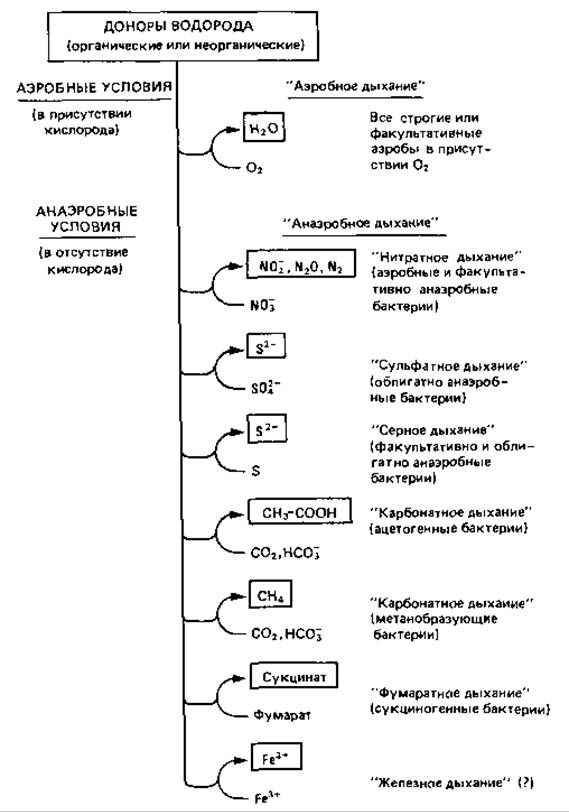
Рис. 9.1. Процессы получения энергии путем окислительного фосфорилирования в аэробных и анаэробных условиях («аэробное дыхание» и «анаэробное дыхание»)
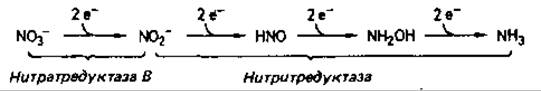
Ассимилиция нитрата. В процессе ассимиляции нитрат сначала восстанавливается до нитрита, а затем до аммиака; последний используется для синтеза аминокислот и других азотсодержащих компонентов клетки. Первый этап катализирует нитратредуктаза; этот фермент (нитратредуктаза В) находится в цитоплазме, и его синтез индуцируется в том случае, если нитрат оказывается единственным источником азота в питательной среде. Нитрит восстанавливается до аммиака с помощью нитритредуктазы, на что затрачивается 6 электронов. Электроны поступают от NAD(P)H2(у грибов и бактерий) или ферредоксина (у растений и некоторых бактерий). Нитритредуктаза - сложный фермент, его каталитические центры содержат атомы железа, из которых одни входят в состав гема (сирогем), а другие связаны с атомами серы. Нитритредуктазная реакция сходна с превращениями, катализируемыми нитрогеназой и сульфитредуктазой, так как при восстановлении субстрата происходит перенос 6 электронов и не высвобождается никаких промежуточных продуктов:
Образование аммония из нитрата представляет собой процесс восстановления, протекающий с большими затратами восстановительной силы:
![]()
Сравнивая рост клеток в аэробных условиях на средах, содержащих в качестве источника азота нитрат или соответственно аммоний, можно заметить, что в первом случае затрачивается больше восстановительной силы (донора водорода), чем при восстановлении аммония (при одинаковом выходе биомассы).
Нитратное дыхание: денитрификация. Денитрифицирующие бактерии обладают способностью восстанавливать нитрат через нитрит до газообразной закиси азота (N2О) и азота (N2):
![]()
В отсутствие кислорода нитрат, таким образом, служит конечным акцептором водорода.
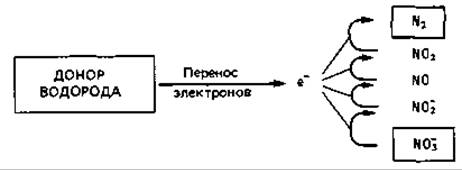
Способность получать энергию путем использования нитрата как конечного акцептора водорода с образованием молекулярного азота широко распространена у бактерий. Этот процесс денитрификации до сих пор был обнаружен только у факультативных аэробов; по-видимому, среди облигатных анаэробов нет денитрифицирующих форм. Кроме того, бактерии чаще обладают полной дыхательной системой; синтез ферментов, необходимых для денитрификации (мембраносвязанные нитратредуктаза А и нитритредуктаза), индуцируется только в анаэробных условиях (рис. 9.2). У многих денитрификаторов эта индукция происходит лишь в присутствии нитрата, хотя для некоторых достаточно создания анаэробных условий. Многие денитрификаторы могут расти, используя в качестве акцептора водорода не только нитрат, но и нитрит, а иногда даже закись азота. Из этого следует, что не только нитратредуктаза А, но и диссимиляционная нитритредуктаза связана у них с дыхательной цепью и участвует в выработке энергии.
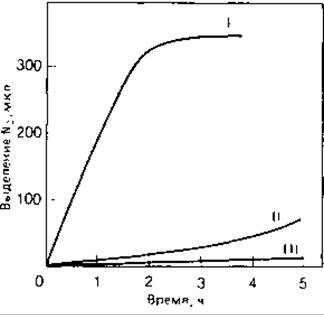
Рис. 9.2. Скорость образования молекулярного азота в клеточной суспензии. N2 образуется в результате денитрификации нитрата в анаэробных условиях (окисляемым субстратом служит ацетат). Бактерии росли в различных условиях: анаэробно в присутствии нитрата (І); аэробно в присутствии нитрата (ІІ); аэробно без нитрата (ІІІ). (Van Olden Е. In: Kluyver A.J., Van Niel С. В. The Microbe’s Contribution to Biology, Cambridge [Mass.]: Harvard Univ. Press, 1956.)
Накопительная культура денитрифицирующих бактерий. Если в среду, содержащую доноры водорода, субстраты для построения клеточных компонентов и нитрат, внести почву или ил и инкубировать культуру без доступа воздуха (см. табл. 6.3), то в ней будут расти различные бактерии: а) при наличии следов пептона и этанола или пропионовой кислоты - Pseudomonas aeruginosa; б) на глюкозе - Pseudomonas fluorescens; в) на тартрате, сукцинате или малате - Pseudomonas stutzeri; г) на среде с органическими кислотами, спиртом, мясным экстрактом и при высокой концентрации нитрата (5-12% KNO3) - Bacillus licheniformis; д) при наличии следов дрожжевого экстракта и Н2 в качестве донора водорода - Paracoccus denitrificans; е) в присутствии серы или тиосульфата - Thiobacillus denitrificans. Поскольку некоторые денитрификаторы используют нитрат исключительно или преимущественно как акцептор водорода, но не могут восстанавливать его до NH4, необходимо добавлять в среду источник азота (пептон или соль аммония).
Обеднение почвы азотом вследствие денитрификации. Временные потери азота на ограниченных участках почвы, несомненно, связаны с деятельностью денитрифицирующих бактерий. Она имеет большое значение, когда в почве создаются анаэробные условия, например при застойном переувлажнении, особенно если при этом применяются органические удобрения и нитраты. На рисовых полях удобрение нитратами может приводить к вредным последствиям из-за накопления нитритов. Нитрит аккумулируется также в содержащих нитрат сточных водах при недостаточном доступе воздуха, а иногда попадает и в источники питьевой воды. Зависимость обеднения почвы азотом от аэрации связана с особенностями регуляции нитратредуцирующей ферментной системы у бактерий. Эти ферменты индуцируются нитратом только в анаэробных условиях (рис. 9.2); молекулярный кислород подавляет (репрессирует) синтез нитрат- и нитритредуктаз. В том случае, если ферменты уже были синтезированы до того, как клетки пришли в соприкосновение с кислородом воздуха, кислород вступает в конкуренцию с нитратом за электроны, поступающие из дыхательной цепи, а также подавляет функцию нитратредуцирующей системы.
Значение денитрификации в природе. Денитрификация-единственный биологический процесс, благодаря которому связанный азот преобразуется в свободный N2. С глобальной точки зрения этот процесс имеет решающее значение для сохранения жизни на земной суше. В нормально аэрируемых почвах и водоемах нитрат представляет собой конечный продукт минерализации. Благодаря своей высокой растворимости в воде и слабому связыванию почвой нитрат-ионы вымывались бы из почвы и накапливались в морской воде; содержание молекулярного азота в атмосфере стало бы уменьшаться, и процессы роста растений и продукции биомассы на суше в конце концов прекратились бы.
Нитратное дыхание: восстановление нитрата до нитрита. Для целого ряда факультативно-анаэробных бактерий (Enterobacter, Escherichia coli и др.) нитрат может служить конечным акцептором водорода в процессе транспорта электронов, поставляющем энергию. Этот вид «нитратного дыхания» отличается от денитрификации тем, что здесь только первая ступень, а именно восстановление нитрата до нитрита с помощью нитратредуктазы А, сопряжена с переносом электронов и преобразованием энергии:
![]()
При этом нитрит может накапливаться в культуральной жидкости - образования N2не происходит.
Однако вместо этого нитрит может восстанавливаться до аммиака путем ассимиляционной нитритредукции с последующим выделением NH4 в среду. В таком случае говорят об аммонификации нитрата. Восстановление нитрита до аммиака не позволяет клетке получать энергию. Речь идет скорее о процессе брожения, в котором нитрит играет роль экзогенного акцептора электронов. Таким образом, восстановление нитрита все же дает известное преимущество: при сбраживании глюкозы часть атомов водорода расходуется на восстановление нитрита, в результате чего образуется больше ацетата (см. стр. 264, реакция 3).
Образование метгемоглобина как следствие накопления нитрата в питьевой воде и пищевых продуктах. Употребление содержащих нитрат питьевой воды (довольно частое в засушливые периоды) и овощей (например, «переудобренного» шпината) может вследствие бактериального восстановления нитрата в консервах или в кишечнике приводить к заболеваниям. Восстановление нитрата до нитрита микроорганизмами с последующим поступлением нитрита в кровь служит причиной образования метгемоглобина, в котором кислород прочно (необратимо) связан с гемоглобином. В результате эритроциты частично утрачивают способность переносить кислород, что приводит к цианозу. Такое заболевание встречается только у детей младше 6 месяцев. У них нитратредуцирующие бактерии, по-видимому, не разрушаются в желудке и попадают в кишечник, где восстанавливают нитрат, растут и накапливают нитрит. У детей более позднего возраста и у взрослых такие бактерии погибают под действием сильнокислого желудочного сока, а ионы нитрата всасываются раньше, чем они могли бы подвергнуться восстановлению при благоприятных для этого значениях pH в двенадцатиперстной кишке.