Общая микробиология - Шлегель Г. 1987
Использование неорганических доноров водорода: аэробные хемолитотрофные бактерии
Фиксация СО2
Большинство организмов, способных расти в условиях, когда единственным источником углерода служит углекислота, фиксируют ее через рибулозобисфосфатный цикл (цикл Кальвина-Бассама). К таким организмам относятся аэробные хемолитоавтотрофные бактерии, почти все фототрофные бактерии, цианобактерии и зеленые растения. Цикл Кальвина-Бассама определенно не участвует в ассимиляции СО2 у метанобразующих и ацетогенных бактерий, хотя они тоже относятся к хемолитоавтотрофным организмам.
Для рибулозобисфосфатного цикла характерны два фермента, не участвующие в других метаболических путях, -фосфорибулокиназа и рибулозобисфосфат-карбоксилаза. Последний фермент представляет собой белок, в количественном отношении преобладающий над всеми другими белками на нашей планете. Рибулозобисфосфатный цикл - это восстановительный процесс, в котором СО2 восстанавливается до уровня углеводов. В цикле могут быть выделены три участка: 1) реакция карбоксилирования, 2) восстановление и 3) регенерация молекул, служащих акцепторами СО2.
Реакция карбоксилирования. При участии рибулозобисфосфат-карбоксилазы к рибулозо-1,5-бисфоефату присоединяется СО2, и в результате образуются две молекулы 3-фосфоглицериновой кислоты:

Этот же фремент может катализировать и другую реакцию. В отсутствие СО2 и в присутствии О2 он проявляет оксигеназную активность, окисляя рибулозобисфосфат до фосфогликолата и 3-фосфоглицерата. Эта реакция участвует в образовании гликолевой кислоты у автотрофных бактерий и у зеленых растений, а следовательно - в фотодыхании.
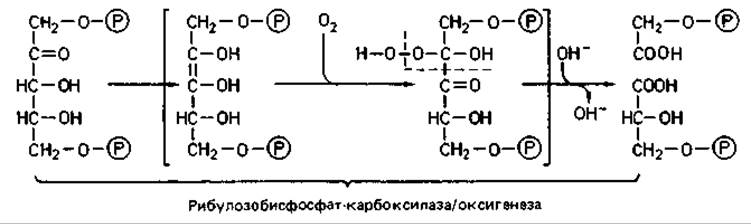
Реакция восстановления. За реакцией карбоксилирования следует восстановление карбоксильной группы 3-фосфоглицерата до альдегидной группы. В нем участвуют реакции, известные для фруктозобисфосфатного пути (разд. 7.2.1), т. е. фосфорилирование под действием 3-фосфоглицераткиназы за счет АТР и восстановление при участии глицеральдегид-3-фосфатдегидрогеназы за счет NAD(P)H2. У бактерий эта реакция зависит от NAD, а у растений-от NADP1.
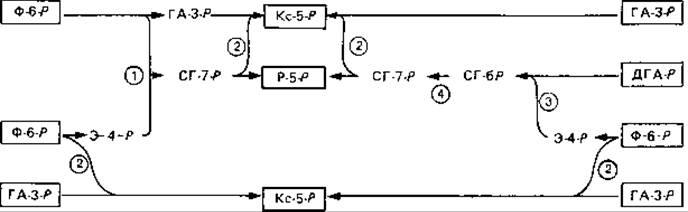
Рис. 11.2. Схема двух возможных путей регенерации пентозофосфатов из триозофосфатов и фруктозо-6-фосфата. Слева - этапы синтеза пентоз нуклеиновых кислот (рибозы, дезоксирибози), протекающие при участии трансальдолазы. Справа - этапы регенерации акцептора СО2 при фиксации СО2 (рибулозо-1,5-бисфосфата), в которых участвуют седогептулозо-1,7-бисфосфат и альдолаза, но уже не участвует трансальдолаза. Ф-6-Р-фруктозо-6-фосфат; ГА-3-Р-глицеральдетид-3-фосфат; ДГА-Р-дигидроксиацетонфосфат; Э-4-Р-эритрозо-4-фосфат; СГ-7-Р-седогептулозо-7-фосфат; СГ-бР-седогептулозо-1,7-бисфосфат; Kc-5-P-ксилулозо-5-фосфат; Р-5-Р-рибозо-5-фосфат. Ферменты (цифры в кружках): I - трансальдолаза; 2-транскетолаза; 3-фруктозобисфосфат-альдолаза; 4 - фруктозобисфосфатаза.
Восстановление 3-фосфоглицерата-это, собственно, и есть тот этап ассимиляции СО2, который требует затраты энергии и восстановительной силы. Последующие стадии происходят на примерно одинаковом энергетическом уровне.
Регенерация акцепторов СО. Глицеральдегид-3-фосфат находится в равновесии с дигидроксиацетонфосфатом (триозофосфатизомеразная реакция), а оба триозофосфата - в равновесии с фруктозо-1,6-бисфосфатом (альдолазная реакция).
Фруктозобисфосфаг дефосфорилируется под действием фруктозобисфосфатазы с образованием фруктозо-6-фосфата. Затем из одной молекулы фруктозо-6-фосфата и трех молекул триозофосфата образуются три молекулы рибозо-5-фосфата. Некоторые из ферментов, катализирующих эти превращения, участвуют и в окислительном пентозофосфатном цикле. Первое из этих превращений - транскетолазная реакция (рис. 11.2, справа). Транскетолаза катализирует перенос гликольной группы от кетозомонофосфата на альдозофосфат. Гликолевый альдегид при этом временно связывается с тиаминдифосфатом (тиаминпирофосфатом), играющим роль кофермента: получается «активный гликолевый альдегид. Образующийся в ходе транскетолазной реакции тетрозофосфат (эритрозо-4-фосфат) превращается в альдолазной реакции с дигидроксиацетонфосфатом в седогептулозо-1,7-бисфосфат. Последний подвергается при участии фруктозобисфофатазы дефосфорилированию в положении 1 с образованием седогептулозо-7-фосфата. Эта реакция гидролиза фосфорного эфира необратима и дает возможность регулировать метаболизм в данном пункте.
1 У цианобактерий в этой реакции восстановления тоже участвует NADPH. Прим. ред.
Для высших растений точно установлено, что в ходе фотосинтеза регенерация рибулозо-5-фосфата происходит через седогептулозо-1,7-бисфосфат (рис. 11.2, справа); однако в темноте синтез пентозофосфатов ведет через трансальдолазную реакцию прямо к седогептулозо-7-фосфату (рис. 11.2, слева).
Гликолильная группа седогептулозо-7-фосфата переносится с помощью транскетолазы на глицеральдегид-3-фосфат, что ведет к образованию двух пентозофосфатов. Эти пентозофосфаты (рибозо-5-фосфат и ксилулозо-5-фосфат) находятся в равновесии с рибулозо-5-фосфатом. Последней реакцией рибулозобисфосфатного цикла является фосфорилирование рибулозо-5-фосфата за счет АТР при участии фосфорибулокиназы до рибулозо-1,5-бисфосфата.
Баланс рибулозобисфосфатного цикла. Для синтеза 1 моля гексозы из 6 молей С02 необходимы шесть оборотов. Баланс фиксации СО2 в рибулозобисфосфатном цикле можно представить следующим уравнением:
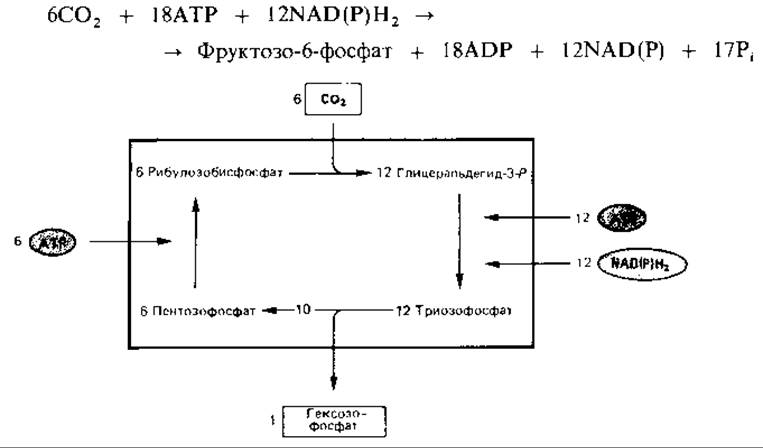
Хотя цикл и изображают замкнутым, многие его промежуточные продукты служат важными исходными веществами для синтеза клеточных компонентов: из 3-фосфоглицерата образуются пируват и ацетил-СоА, из эритрозо-4-фосфата - ароматические аминокислоты; рибозо-5-фосфат используется для синтеза нуклеотидов, а гексозофосфаты - для построения полимеров. Регуляция активности некоторых участвующих в цикле ферментов преследует, видимо, две цели: с одной стороны, на фиксацию СО2 (связанную с большой затратой энергии) не должно расходоваться слишком много АТР; с другой стороны, цикл не должен прерваться (что могло бы произойти, если бы его промежуточные продукты оказались использованными; стр. 496-497).
Другие пути автотрофной фиксации СО2. Фиксация СО2 в рибулозобисфосфатном цикле является ныне хотя и важнейшей для биосферы, но далеко не единственной цепью реакций, ведущей к синтезу органических веществ. Анаэробные автотрофные бактерии располагают двумя другими механизмами ассимиляции СО2. Метанобразующие, ацетогенные и сульфатредуцирующие (сульфидогенные) бактерии, способные использовать в качестве донора электронов Н2 или СО, восстанавливают СО2 по анаэробному ацетил-СоА-пути до ацетил-СоА и пирувата (разд. 9.4). Последний вступает в результате известных реакций на центральные пути биосинтеза.
Зеленые серобактерии (Chlorobium limicola, forma thiosulfatophilum) фиксируют СО2 исключительно с помощью реакций восстановительного цикла трикарбоновых кислот; СО2 фиксируется благодаря восстановительному карбоксилированию сукцинил-СоА.
Сравнение всех трех типов автотрофной фиксации СО2 позволяет заключить, что анаэробные процессы более экономны, чем аэробные. Синтез 1 моля триозофосфата из 3 молей СО2 по анаэробному ацетил-СоА-пути требует затраты всего лишь 3 молей АТР, в восстановительном цикле трикарбоновых кислот-затраты 5 молей АТР, а в рибулозобисфосфатном цикле - 9 молей АТР. Общие реакции фиксации СО2. Мы уже не раз упоминали о том, что и гетеротрофные организмы нуждаются в двуокиси углерода и вовлекают ее в свой метаболизм. Отмечалась роль карбоксилирования пирувата и фосфоенолпирувата в функционировании цикла трикарбоновых кислот. В какие промежуточные продукты обмена может включиться СО2, показано на рис. 11.3.
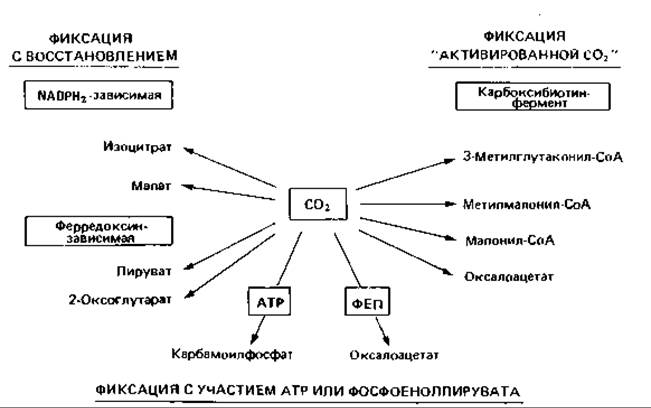
Рис. 11.3. Включение СО2 в различные промежуточные продукты метаболизма.
Упоминавшиеся реакции фиксации СО2 играют различную роль у разных организмов. Некоторые из них служат для активации метаболитов или для пополнения центральных биохимических путей метаболитами. Реакции восстановительного карбоксилирования, зависимые от ферредоксина, встречаются лишь у некоторых анаэробных и фототрофных бактерий.