Общая микробиология - Шлегель Г. 1987
Разложение природных веществ
Целлюлоза
Хотя зеленые растения на протяжении многих миллионов лет синтезируют из двуокиси углерода органические соединения, сколько-нибудь заметного накопления органических веществ за это время не произошло. Лишь небольшая их часть в условиях без доступа воздуха сохранилась в форме сильно восстановленных соединений углерода; это нефть, природный газ и каменный уголь. В аэробных условиях все вещества биологического происхождения подвергаются распаду. Каким бы сложным ни было то или иное вещество, в природе всегда найдется микроорганизм, способный полностью или частично его расщепить, а продукты этого расщепления будут использованы другими микроорганизмами. Таким образом, в совокупности микроорганизмы в биохимическом смысле «всемогущи», и это дает основание говорить об «универсальности» микробов. В настоящее время, однако, нужно внести в это утверждение некоторые коррективы. Многие из созданных человеком низкомолекулярных веществ (ядохимикаты, детергенты и т. п.) и высокомолекулярных полимеров оказались устойчивыми и не разлагаются микроорганизмами (насколько позволяют судить многолетние наблюдения и результаты экспериментов).
Сведения о микроорганизмах, которые воздействуют на отдельные природные вещества, разлагая и перерабатывая их, получены главным образом в опытах с накопительными культурами. Простые питательные среды, содержащие в качестве источника энергии исследуемое природное вещество, обеспечивают рост только тех микроорганизмов, у которых потребности в питательных веществах не слишком сложны; кроме того, в жидких накопительных культурах выживают лишь наиболее быстро растущие в данных условиях организмы. Поэтому возникает сомнение, можно ли подобные формы считать типичными представителями организмов, осуществляющих те же процессы разложения в природе. При такой постановке опытов многие виды, для роста которых нужны иные условия, остаются невыявленными. В наших знаниях о разложении природных веществ еще много пробелов, а биохимический анализ путей этого разложения только начинается.
Целлюлоза является главной составной частью всякого растительного материала, и синтез ее по своим масштабам превосходит синтез всех других природных соединений. Сохраняющиеся в почве и возвращающиеся в нее растительные остатки на 40-70% состоят из целлюлозы. Столь большое количество целлюлозы в природе обусловливает важную роль разлагающих ее микроорганизмов в процессах минерализации и в круговороте углерода (разд. 1.3).
Целлюлоза состоит из цепочек ß-D-глюкозы со степенью полимеризации около 14000 (разд. 2.2.3). Физические свойства целлюлозных фибрилл (особенно их механическая прочность и нерастворимость) зависят не от структуры отдельных цепочек. Цепочки должны быть связаны между собой таким образом, чтобы гидрофильные группы были скрыты (это повышает стабильность). По данным рентгеноструктурного анализа, участки, имеющие кристаллическое строение, чередуются в целлюлозе с некристаллическими участками. Целлюлозные волокна представляют собой пучки фибрилл, одетые общей оболочкой, которая содержит воск и пектин.
Ферментативное расщепление целлюлозы осуществляется под воздействием целлюлазы. В экспериментах на грибах было показано, что в систему целлюлазы входят по меньшей мере три фермента: 1) эндо-ß-1,4-глюканаза одновременно разрывает различные ß-1,4-связи внутри макромолекулы, что ведет к образованию больших фрагментов со свободными концами; 2) экзо-β-1,4-глюканаза отщепляет от конца цепочки дисахарид целлобиозу; 3) ß-глюкозидаза осуществляет гидролиз целлобиозы с образованием глюкозы.
В условиях лабораторной культуры микроорганизмы обычно синтезируют эти ферменты лишь в том случае, если целлюлоза -единственный имеющийся субстрат. Их синтез подавляют как другие субстраты, так и продукт расщепления целлюлозы - целлобиоза.
Разложение целлюлозы в аэробных условиях. В хорошо аэрируемых почвах целлюлозу разлагают и используют аэробные микроорганизмы (грибы, миксобактерии и другие эубактерии), а в анаэробных условиях - в основном клостридии.
В аэробных условиях значительная роль в разложении целлюлозы принадлежит грибам. Они в этом отношении эффективнее бактерий, особенно в кислых почвах и при разложении целлюлозы, инкрустированной лигнином (древесины). Большую роль играют в этом процессе представители двух родов - Fusarium и Chaetomium. Целлюлозу расщепляют также Aspergillus fumigatus, А. nidulans, Botrytis cinerea, Rhizoctonia solani, Trichoderma viride, Chaetomium globosum и Myrothecium verrucaria. Три последних вида служат тест-организмами для выявления распада целлюлозы, а также при испытании средств, применяемых для пропитки различных материалов с целью предохранить их от действия микроорганизмов, разлагающих целлюлозу. Грибы образуют целлюлазы, которые можно выделить из мицелия и из питательной среды.
Cytophaga и Sporocytophaga аэробные бактерии, разлагающие целлюлозу. Их легче всего выделить обычным методом накопительной культуры в жидких средах. Эти два рода, близкие к миксобактериям, включают много видов. Об использовании целлюлозы миксобактериями и об их первичном воздействии на нее мало что известно. У них не удалось обнаружить ни внеклеточной целлюлазы, ни каких-либо продуктов расщепления целлюлозы. Клетки этих бактерий тесно прилегают к волокнам целлюлозы, располагаясь параллельно оси волокна. По-видимому, они гидролизуют целлюлозу лишь при тесном контакте с волокном, и продукты гидролиза тотчас же поглощаются. На агаре с целлюлозой колонии Cyiophaga никогда не бывают окружены прозрачной зоной, в которой находились бы продукты ферментативного расщепления целлюлозы.
Кроме видов Cytophaga на целлюлозе могут расти миксобактерии родов Polyangium, Sporangium и Archangium, образующие плодовые тела.
Использовать целлюлозу как субстрат для роста могут и многие из тех аэробных бактерий, которые можно было бы назвать «всеядными». Некоторые из них используют целлюлозу, видимо, только в тех случаях, когда нет других источников углерода; синтез и выделение целлюлаз у таких бактерий регулируются по типу катаболитной репрессии. Некоторые формы, сходные с Pseudomonas, раньше объединяли в группу Cellvibrio. Сейчас их описывают как Pseudomonas fluorescens var. celłułosa. Из коринеформных бактерий следует упомянуть Cellulomonas; эту бактерию предполагали даже использовать для получения белка из целлюлозы.
Среди актиномицетов описано лишь немного целлюлозоразлагающих видов: Micromonospora chalcea, Streptomyces cellulosae, Streptosporangium.
Разложение целлюлозы в анаэробных условиях. В анаэробных условиях целлюлозу расщепляют чаще всего мезофильные и термофильные клостридии. Термофильный вид Clostridium thermocellum растет на простых синтетических средах, используя в качестве субстрата целлюлозу или целлобиозу, а в качестве источника азота - соли аммония; глюкозу и многие другие сахара эта бактерия не утилизирует. Продуктами сбраживания целлюлозы являются этанол, уксусная, муравьиная и молочная кислоты, молекулярный водород и СО2. Вне клеток целлюлоза расщепляется, вероятно, только до целлобиозы. К сходным продуктам приводит сбраживание целлюлозы мезофильным видом Clostridium cellobioparum. Длинная палочка Bacillus dissolvens ведет себя подобно упомянутым выше видам Cytophaga: клетки этой бактерии тесно прилегают к волокнам целлюлозы и не выделяют в среду целлюлазы.
Микробиологические процессы в рубце жвачных животных. В рубце жвачных целлюлозу тоже расщепляют главным образом бактерии. Основными источниками углеводов для жвачных служат сено, солома и трава. В сухой траве примерно половину углеводов составляют фруктозаны и ксиланы и приблизительно столько же приходится на долю целлюлозы. Целлюлозные компоненты кормов были бы недоступны для использования, если бы у жвачных в процессе эволюции не возникли симбиотические отношения с микробами, способными расщеплять целлюлозу (рис. 14.1).
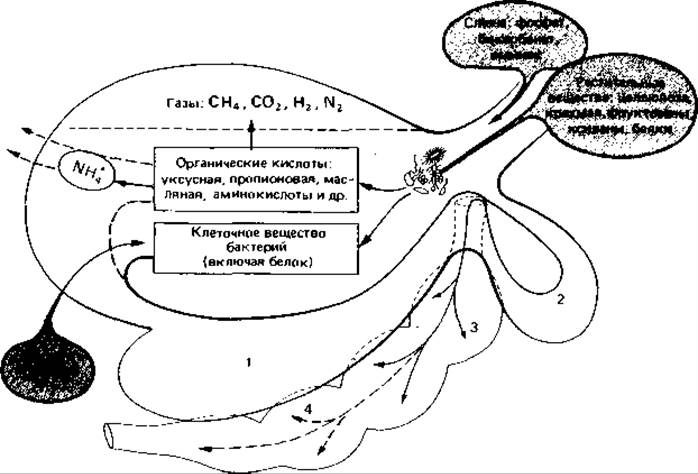
Рис. 14.1. Схема превращений, которым подвергаются питательные вещества под действием микроорганизмов в желудке жвачных. 1 - рубец; 2 - сетка; 3 - книжка; 4 - сычуг. Объяснение см. в тексте.
Два первых отдела желудка жвачных - рубец и сетка - это как бы большая бродильная камера (емкостью от 100 до 250 л), в которой существуют идеальные условия для роста многочисленных микроорганизмов; им обеспечены здесь постоянная температура (37-39°С), непрерывная подача минерального раствора (около 100-200 л слюны в сутки), хорошо забуференного бикарбонатом и фосфатом (pH 5,8-7,3), периодическое поступление питательных веществ в виде хорошо размельченного, богатого целлюлозой корма и, наконец, механическое перемешивание в результате движений рубца. Таким образом, рубец напоминает систему для полунепрерывного культивирования микроорганизмов.
Среди обитателей рубца преобладают простейшие и бактерии. В 1 мл содержимого рубца находится несколько миллионов простейших- главным образом инфузорий, относящихся к родам Diplodinium и Entodinium. Это специфические для рубца, мало распространенные в других местах виды. По массе они составляют от 6 до 10% содержимого рубца, причем часть этой массы приходится на долю запасенных ими полисахаридов. Однако простейшие, возможно, не играют в рубце жизненно важной роли. Неясно, участвуют ли они в разложении целлюлозы.
С функциональной точки зрения наиболее важными обитателями рубца являются бактерии. В 1 мл рубцовой жидкости содержится от 109 до 1010 бактериальных клеток. На их долю приходится 5-10% сухой массы содержимого рубца. Специфические для рубца бактерии — строгие анаэробы. Дрожжи и другие грибы присутствуют здесь лишь в небольшом количестве.
Бактерии перерабатывают полимерные углеводы кормов в простые соединения, такие как жирные кислоты и спирты. Из целлюлозы, крахмала, фруктозана и ксилана образуются в основном жирные кислоты. По имеющимся данным, разложению подвергается около 90% (по весу) всей поступающей с кормом целлюлозы. При этом образуются большие количества кислот, главным образом уксусной (50-70 об. %), пропионовой (17-21 об. %) и масляной (14 -20 об. %), а также немного валериановой и муравьиной. Кроме того, ежедневно образуется до 900 л газа примерно следующего состава (по объему): 65% СО2, 27% метана, 7% N2, 0,18% Н2 и следовые количества сероводорода. Сравнительно недавно из содержимого рубца были выделены бактерии, которые и в лабораторных условиях сбраживали целлюлозу до тех же кислот и в тех же соотношениях, что и в рубце. Поэтому можно предполагать, что органические кислоты в рубце образуются в результате разложения целлюлозы бактериями.
Расщеплять целлюлозу в рубце способны Ruminococcus albus и R. flavefaciens - грам-отрицательные кокки; Bacteroides succinogenes - грам-отрицательная неподвижная палочка, образующая главным образом уксусную и янтарную кислоты; Butyrivibrio fibrisolvens; Clostridium cellobioparum.
Отсутствие в рубце молочной кислоты следует приписать активности бактерии Veillonella alcalescens (Micrococcus lactilyticus), сбраживающей лактат с образованием пропионата, ацетата, молекулярного водорода и СО2. Метан не является прямым продуктом разложения целлюлозы: он имеет вторичное происхождение и образуется из жирных кислот, а также молекулярного водорода и СО2 (разд. 9.4). Образование сероводорода в рубце связано с восстановлением сульфата бактерией Desulfotomaculum ruminis. Selenomonas ruminantium (рис. 2.36, Б) сбраживает глюкозу в молочную, уксусную и пропионовую кислоты.
В местах естественного обитания жвачных - в саваннах и степях - их корм очень беден азотом и белками. Синтез белка обеспечивает симбиотическая микрофлора рубца. Дело в том, что у жвачных сформировался весьма эффективный «желудочно-печеночный цикл». Мочевина, образующаяся в печени в процессе обезвреживания аммиака, лишь частично выводится с мочой; вся остальная мочевина поступает через слюнные железы и стенку рубца в первые отделы желудка и может использоваться микроорганизмами рубца для синтеза белка (рис. 14.1). Благодаря симбиотическим взаимоотношениям с микроорганизмами рубца жвачные не зависят от экзогенных источников белка. Было неоднократно показано, что коров можно держать на безбелковом корме.
Бактерии играют в питании жвачных двоякую роль. Кислоты, образующиеся при распаде полисахаридов, всасываются здесь же в рубце. Сами бактерии при переходе содержимого рубца в кишечник перевариваются, так что вещество их клеток тоже подвергается разложению и усваивается животным. Поскольку бактерии рубца используют и не органические источники азота, это существенно увеличивает количество белка, получаемое организмом.
Бактерии рубца подвергают растительные жиры гидрированию. Образующиеся насыщенные жирные кислоты всасываются в кишечнике, а затем включаются в собственные жиры крупного рогатого скота, входящие в состав мяса, молока и масла. У животных, не имеющих рубца, подобного повышения тугоплавкости жира не происходит. Жиры, накапливаемые в организме свиньи или грызунов, имеют поэтому более мягкую констистенцию (более низкую температуру плавления), чем жиры жвачных; они содержат ненасыщенные жирные кислоты и кислоты с более короткой цепью, т. е. те, которые поступают с растительным кормом. Если учесть, что не только жиры изменяются под влиянием бактерий рубца, но и 60-90% (по весу) белков крупного рогатого скота имеет бактериальное происхождение, то можно, поедая говяжий бифштекс, поистине чувствовать себя в гостях у бактерий, а наслаждаясь свиным шницелем - в гостях у кормовых растений!