ЗАГАЛЬНА ГІСТОЛОГІЯ З ОСНОВАМИ ЕМБРІОЛОГІЇ - Долгов О. М. - 2015
Частина І
БІОЛОГІЯ КЛІТИНИ
КЛІТИННИЙ ЦИКЛ
Одне з положень клітинної теорії проголошує, що збільшення кількості клітин, їх розмноження, відбувається шляхом поділу вихідної клітини. Воно повністю заперечує будь-яке «самозародження» клітин або їх утворення із неклітинної «живої речовини». Зазвичай поділу клітин передує редуплікація їхнього хромосомного апарату, синтез ДНК. Це правило є загальним для прокаріотичних і еукаріотичних клітин. Функція відтворення і передачі генетичної інформації забезпечується в ході клітинного циклу.
Клітинний цикл - час існування клітини як такої - від поділу до поділу або від поділу до загибелі. Його тривалість може бути різною для різних типів клітин. Так, наприклад, для бактеріальних клітин у стаціонарних умовах культивування цей час може складати 20-30 хвилин. У еукаріотичних одноклітинних організмів тривалість життя клітини значно довша. Інфузорія туфелька може поділятися 1-2 рази на добу, час клітинного циклу при безстатевому розмноженні у амеби складає 1,5 доби, у інфузорії трубача - 2-3 доби. Тривалість клітинного циклу залежить від температури та умов навколишнього середовища.
Клітини багатоклітинних організмів мають різну здатність до поділу. Якщо у ранньому ембріогенезі клітини тварин поділяються часто, то у дорослому організмі вони в більшості втрачають цю здатність. У круглих червів і коловраток клітини втрачають здатність до поділу після закінчення ембріогенезу, і ріст організму, наприклад у аскариди, відбувається не за рахунок збільшення числа клітин, а за рахунок збільшення їхніх розмірів.
Клітинний цикл включає власне мітотичний поділ і інтерфазу - проміжок між поділами (Рис. 25).
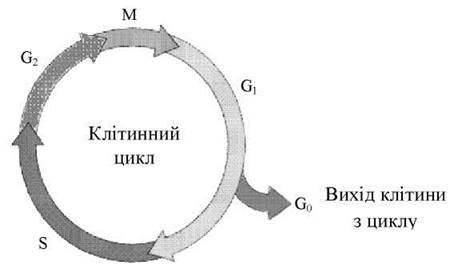
Рис. 25. Схема клітинного циклу. Пояснення в тексті.
Інтерфаза значно більш тривала, ніж мітоз (звичайно займає не менш 90% усього часу клітинного циклу) і підрозділяється на три періоди: передсинтетичний, або післямітотичний (G1), синтетичний (S) і післясинтетичний, або передмітотичний G2).
Передсинтетичний, або післямітотичний (G1-) період (англ. gap - проміжок) настає відразу ж після мітотичного поділу і характеризується активним ростом клітини та синтезом білка і РНК, завдяки чому клітина досягає відповідних розмірів і відновлює необхідний набір органел. G1-період триває від декількох годин до декількох днів. Протягом цього періоду синтезуються особливі білки «запуску» (trigger proteins), або активатори S-періоду. Вони забезпечують досягнення клітиною визначеного порога (точки R - рестрикції або обмеження), після якого вона вступає в S-період.
Контроль, здійснюваний на рівні точки R (при переході з G1 у S), обмежує можливість нерегульованого розмноження клітин. Проходячи цю точку, клітина переключається на наступну регуляцію внутрішніми факторами клітинного циклу, що забезпечує закономірне завершення її поділу. Якщо клітина не досягає точки R, вона виходить з циклу і вступає в період репродуктивного спокою (G0) для того, щоб (залежно від причин зупинки): (1) диференціюватися і виконувати свої специфічної функції, (2) вижити в умовах недостатності поживних речовин або факторів росту, (3) здійснити репарацію ушкодженої ДНК. Клітини одних тканин при відповідній стимуляції знову здатні повертатися з періоду (G0) у клітинний цикл, інших - втрачають цю здатність у міру диференціювання.
Синтетичний (S-) період характеризується подвоєнням кількості (реплікацією) ДНК і синтезом білків, зокрема, гістонів, що надходять у ядро з цитоплазми і забезпечують нуклеосомне упакування знову синтезованої ДНК. У результаті відбувається подвоєння числа хромосом. Одночасно подвоюється число центріолей. S-період триває в більшості клітин 8-12 годин.
Післясинтетичний, або передмітотичний (G2-)період настає за S-періодом і продовжується аж до мітозу (що часто позначається буквою М). Протягом цього періоду клітина здійснює безпосередню підготовку до поділу. Відбувається дозрівання центріолей, запасається енергія у вигляді АТФ, синтезуються РНК і білки (зокрема, тубуліни), необхідні для процесу поділу. Тривалість G2-періодy складає 2-4 години. Можливість виходу клітини з G2-періодy в G0-період з наступним поверненням у G2-період у даний час більшістю авторів заперечується.
Контроль вступу клітини в мітоз здійснюється двома спеціальними факторами з протилежно спрямованими ефектами. Мітоз гальмується до моменту завершення реплікації ДНК М-затримуючим фактором та індукується М-стимулюючим фактором. Дія останнього виявляється лише в присутності інших білків - циклінів (синтезуються протягом усього циклу і розпадаються в середині мітозу).
За рівнем відновлення клітин усі тканини організму підрозділяються на три групи.
Стабільні клітинні популяції складаються з клітин з повною втратою здатності до поділу (переважна кількість нейронів, кардіоміоцити). Кількість клітин у такій популяції стабілізується на початку їхнього диференціювання; у міру старіння організму вона знижується внаслідок природного некомпенсованого збитку клітин.
Зростаючі клітинні популяції здатні не тільки до відновлення, але також і до росту, збільшенню маси тканини за рахунок наростання числа клітин і їх поліплоїдізації. Їх клітини, що живуть тривалий час, виконують спеціалізовані функції, але зберігають здатність при стимуляції знову вступати в цикл для того, щоб відновити свою нормальну чисельність. Описані популяції клітин утворюють нирки, печінку, підшлункову і щитоподібну залози.
Клітинні популяції, що оновлюються, характеризуються постійним відновленням клітин; збиток диференційованих клітин, які виконують спеціалізовані функції і нездатні до поділу, внаслідок їхньої загибелі урівноважений утворенням нових в результаті поділу малодиференційованих камбіальних клітин та їх наступного диференціювання. До таких популяцій відносять епітелій кишки, епідерміс шкіри, а також клітини кісткового мозку і крові.
Регуляція клітинного циклу в різних тканинах організму здійснюється збалансованою складною системою механізмів, що стимулюють або гальмують клітинний поділ. Система регуляції клітинного циклу одержує два види інформації:
1) про дію на клітину різних зовнішніх факторів, що сприяють активації або гальмуванню її поділу. Вона обробляє й інтегрує її у вигляді сигналів, що визначають, чи буде клітина вступати в мітотичний цикл або диференціюватися і перебувати в періоді репродуктивного спокою (G0);
2) про інтактність (незмінність) геному. При ушкодженні геному клітини проходження нею циклу зупиняється і включається система репарації ДНК. Тим самим знижується ймовірність небажаної реплікації ушкодженої ДНК. Численні сигнали, що регулюють діяльність клітини, замикаються на ген р53, що блокує проходження клітинного циклу до усунення виниклого ушкодження. Якщо це ушкодження занадто серйозне, ген р53 (у сукупності з іншими регуляторами) запускає програму апоптозу - запрограмованої загибелі клітини.
Мітоз
Існують три способи поділу клітин: мітоз, амітоз і мейоз1.
Мітоз або каріокінез (непрямий поділ) - універсальний широко розповсюджений спосіб поділу клітин. При цьому деконденсовані і вже редупліковані хромосоми переходять у компактну форму мітотичних хромосом, утворюється веретено поділу, що бере участь у сегрегації і переносі хромосом (ахроматиновий мітотичний апарат), відбувається розходження хромосом до протилежних полюсів клітини і поділ тіла клітини (цитокінез, цитотомія).
Процес непрямого поділу клітин прийнято підрозділяти на кілька основних фаз: профаза, метафаза, анафаза, телофаза (Рис. 26).
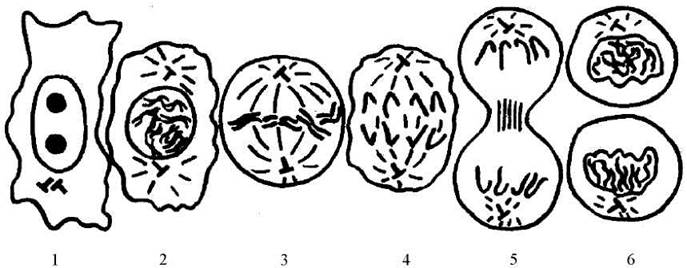
Рис. 26. Схема мітозу.
1 - інтерфаза, 2 - профаза, 3 - метафаза, 4 - анафаза, 5, 6 - телофаза і цитокінез.
Профаза. Після закінчення S-періоду кількість ДНК у інтерфазному ядрі дорівнює 4 с, тому що відбулося подвоєння хромосомного матеріалу. Однак морфологічно реєструвати подвоєння числа хромосом у цій стадії не завжди вдається. Власне хромосоми як нитковидні щільні тільця починають виявлятися мікроскопічно на початку процесу поділу клітини, а саме в профазі мітотичного поділу клітини. Якщо спробувати підрахувати число хромосом у профазі, то їхня кількість буде дорівнювати 2n. Але це помилкове враження, тому що в профазі кожна з хромосом подвійна, що є результатом їхньої редуплікації в інтерфазі. У профазі пари сестринських хромосом тісно стикаються одна з одною, взаємно спіралізуючись одна щодо іншої, тому важко побачити подвійність усієї структури в цілому. Пізніше хромосоми в кожній такій парі починають відокремлюватися, розкручуватися. Така подвійність хромосом у мітозі спостерігається в живих клітинах наприкінці профази, коли видно, що загальне число хромосом у клітині, що почала поділятися, дорівнює 4n. Отже, уже на початку профази хромосоми складалися з двох сестринських хромосом, або, як їх ще називають, хроматид. Число їх (4n) у профазі точно відповідає кількості ДНК (4с).
Паралельно конденсації хромосом у профазі відбувається зникнення, дезинтеграця ядерець у результаті інактивації рибосомних генів у зоні ядерцевих організаторів.
Одночасно з цим у середині профази починається руйнування ядерної оболонки: зникають ядерні пори, оболонка розпадається спочатку на фрагменти, а потім на дрібні мембранні пухирці. Змінюються в цей час і структури, зв’язані із синтезом білка. Відбувається зменшення кількості цистерн гранульованого ендоплазматичного ретикулуму, він розпадається на короткі цистерни і вакуолі, кількість рибосом на його мембранах різко падає. Значно (до 25%) редукується число полісом як на мембранах, так і в гіалоплазмі, що визначає загальне падіння синтезу білка в клітинах, що поділяються.
Друга найважливіша подія при мітозі теж відбувається під час профази - це утворення веретена поділу. У профазі вже репродуковані в S-періоді центріолі починають розходитись до протилежних кінців клітини, де будуть пізніше формуватися полюси веретена. До кожного полюса відходить по подвійній центріолі, диплосомі. В міру розходження диплосом починають формуватися мікротрубочки, що відходять від периферичних ділянок однієї з центріолей кожної диплосоми.
Метафаза. Займає біля третини часу всього мітозу. Під час метафази закінчується утворення веретена поділу, а хромосоми розташовуються в екваторіальній площині веретена, утворюючи так звану метафазну пластинку хромосом, або материнську зірку. До кінця метафази завершується процес відокремлення одна від одної сестринських хроматид. Їхні плечі лежать паралельно одне до іншого, між ними добре видно щілину, що розділяє їх. Останнім місцем, де контакт між хроматидами зберігається, є центромера.
Анафаза. Хромосоми всі одночасно втрачають зв’язок одна з одною в області центромер і синхронно починають віддалятися в напрямку до протилежних полюсів клітини. Швидкість руху хромосом рівномірна, вона може досягати 0,2-0,5 мкм/хв. Анафаза - найкоротша стадія мітозу (кілька відсотків від усього часу), але за цей час відбувається ряд подій. Головною з них є відокремлення двох ідентичних наборів хромосом і переміщення їх у протилежні кінці клітини.
Рух хромосом складається з двох процесів: розходження їх у напрямку до полюсів і додаткового розходження самих полюсів. До того ж, помічено, що клітина, що поділяється, видовжується в напрямку розташування веретена поділу. Оскільки білок тубулін, з якого складаються мікротрубочки, не виявляє АТФазної активності, мікротрубочки не здатні до скорочення. Очевидно, існує механізм їх укорочення в місці прикріплення до хромосоми шляхом дисоціації окремої глобули тубуліну і приєднання кінетохору до наступної.
Телофаза. Починається з зупинки диплоїдних (2n) наборів хромосом, що відокремились, (рання телофаза) і закінчується початком реконструкції нового інтерфазного ядра (пізня телофаза, ранній G1-період) та поділом вихідної клітини на дві дочірні (цитокінез, цитотомія). У ранній телофазі хромосоми, не змінюючи своєї орієнтації (центромерні ділянки - до полюса, теломерні - до екватору), починають деконденсуватися і збільшуватися в об’ємі. У місцях контактів теломерних ділянок хромосом з мембранними пухирцями цитоплазми утворюється нова ядерна оболонка. Після замикання ядерної оболонки починається формування нових ядерець. Клітина переходить у G1-період.
Важлива подія телофази - поділ клітинного тіла, цитотомія, або цитокінез, що відбувається в клітинах тварин шляхом утворення перетяжки, вп’ячування плазматичної мембрани усередину клітини. При цьому в кортикальному, підмембранному шарі цитоплазми розташовуються скоротливі елементи типу актинових фібрил, орієнтовані циркулярно в зоні екватора клітини. Скорочення такого кільця приведе до вп’ячування плазматичної мембрани в області цього кільця, що завершиться поділом клітини на дві.
При ушкодженні мітотичного апарату (дія холоду або агентів, що викликають деполімеризацію тубулінів) може відбутися або затримка мітозу в метафазі, або розсіювання хромосом. При порушеннях репродукції центріолей можуть виникати багатополюсні та асиметричні мітози і т.д. Порушення цитотомії приводить до появи гігантських ядер або багатоядерних клітин.
1 Розглядається в розділі «Основи порівняльної ембріології»