ЗАГАЛЬНА ГІСТОЛОГІЯ З ОСНОВАМИ ЕМБРІОЛОГІЇ - Долгов О. М. - 2015
Частина І
ОСНОВИ ПОРІВНЯЛЬНОЇ ЕМБРІОЛОГІЇ
ГАСТРУЛЯЦІЯ
Гаструляція - складний процес хімічних і морфогенетичних змін, який супроводжується розмноженням, ростом, спрямованим переміщенням і диференціюванням клітин, внаслідок чого утворюються зародкові листки (ектодерма, мезодерма і ентодерма) - джерела зачатків тканин і органів.
На початку гаструляції утворюються зовнішній та внутрішній зародкові листки, а потім на стадії пізньої гаструляції - хордомезодермальний зачаток. Зародкові листки розташовуються пошарово: зовні - ектодерма, всередині - ентодерма, між ними - хорда і мезодерма. По закінченні гаструляції або ще протягом неї виділяються також осьові зачатки органів.
Зародкові листки і осьові зачатки утворюються внаслідок розмноження, переміщення і диференціювання клітинного матеріалу. Диференціювання - процес морфологічної, хімічної і функціональної спеціалізації, яка визначається детермінованістю клітин. Переважно він відбувається шляхом індукції, т.т. впливом зовнішніх факторів і клітин одна на одну. Згідно з теорією організаційних центрів (Х. Шпеман) у певних місцях зародка виникають організуючі фактори, які впливають на інші ділянки зародка, індукуючи їхній розвиток у певному напрямку.
Явище ембріональної індукції було відкрито в 1901 році при вивченні утворення зачатка кришталика ока у зародків земноводних. Гіпотезу про механізм диференціювання, що одержав назву ембріональної індукції, на підставі експериментальних даних висунули Шпеман і Мангольд у 1924 році.
Відповідно до цієї гіпотези, існують певні клітини, що діють як організатори на інші, піддатливі для цього клітини. В умовах відсутності клітин-організаторів такі клітини обирають інший шлях розвитку, відмінний від того, по якому вони розвивалися б в умовах присутності організаторів. Проілюструвати це можна тими самими експериментами 1924-го року, що показали, що диференціювання в значній мірі контролюється впливом цитоплазми клітин одного типу на клітини іншого типу.
Х. Шпеман і його співробітниця Х. Мангольд відкрили в зародків амфібій «організатор». Контрольний експеримент був проведений Хильдою Мангольд у 1921 році. Вона вирізувала шматочок тканини з дорсальної губи бластопора гаструли гребінчастого тритона (Triturus cristatus) зі слабопігменованим зародком, і пересаджувала його у вентральну область іншої гаструли близького виду, тритона звичайного (T. vulgaris), зародок якого характеризується рясною пігментацією. Ця природна різниця в пігментації дозволила розрізнити в химерному зародку тканини донора і реципієнта. Клітки дорсальної губи при нормальному розвитку утворюють хорду і мезодермальні соміти (міотоми). Після пересадження у гаструли-реципієнта з тканин трансплантата розвивалася друга хорда і міотоми. Над ними з ектодерми реципієнта виникала нова додаткова нервова трубка. У підсумку це привело до утворення осьового комплексу органів другого пуголовка на тім же зародку. Ділянка дорсальної губи бластопора, що при пересадженні викликає на новому місці утворення мезодерми і нейроектодерми, одержала назву «організатор Шпемана».
Явище ембріональної індукції тісно зв’язано з такими поняттями, як морфогенез і морфогенетичне поле. Ще Шпеманом було показано, що інактивовані нагріванням тканини організатора зберігають індуктивну активність, і середовище з-під ізольованого організатора також індукує ектодерму.
Пізніше було показано, що багато тканин дорослих тварин індукують нейруляцію ектодерми, також були відкриті речовини- індуктори, такі як хордин і ноггін (діють побічно, через пригнічення BMP (англ. Bone Morphogenetic protein) - епідермального індуктора, його інактивація хордином і ноггіном викликає нейруляцію ектодерми) і багато інших.
Ембріональна індукція - лише один з механізмів онтогенезу. Багатьом явищам розвитку потрібні інші механізми.
За своє відкриття Ханс Шпеман отримав у 1935 р. Нобелівську премію.
Майже у всіх тварин гаструляція здійснюється шляхом інтенсивних морфогенетичних рухів. В залежності від типу бластули і від того який морфогенетичний рух превалює (інколи - лише один) розрізняють 4 основних способи гаструляції (Рис. 46):
Інвагінація (лат. vagina - піхва) - вп’ячування частини стінки бластули (бластодерми) усередину зародка, внаслідок чого утворюється гаструла з порожниною - гастроцелем, який з’єднується із зовнішнім середовищем отвором - бластопором.
Імміграція - виселення у бластоцель окремих клітин бластодерми з одного місця (уніполярна імміграція) або з різних (мультиполярна імміграція).
Епіболія (гр. epibolē - накидання, кладка) - обростання великих малорухомих клітин вегетативного полюсу зародка більш дрібними клітинами його анімальної області.
Деламінація (лат. lamina - пластинка) або розшарування - ентодерма утворюється або шляхом тангенціального (паралельно поверхні) поділу клітин, або шляхом диференціювання первинно однорідних клітин морули (без їхнього поділу) на екто- і ентодерму.
Зазвичай гаструляція здійснюється поєднанням різних її способів.
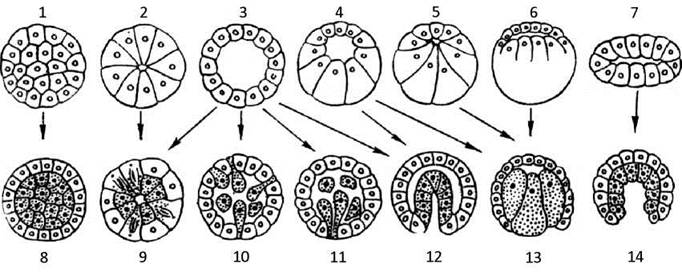
Рис. 46. Типи бластул і характерні для них способи гаструляції:
1 - симетрична морула, 2 - симетрична стерробластула, 3 - симетрична целобластула, 4 - асиметрична целобластула, 5 - асиметрична стерробластула, 6 - дискобластула, 7 - плакула, 8 - морульна деламинація, 9 - клітинна деламінація, 10 - мультиполярна імміграція, 11 - уніполярна імміграція, 12 - інвагінація, 13 - епіболія, 14 - згинання плакули.
Мезодерма утворюється або незалежно від первинних зародкових листків, або первісно входить до складу одного з них і відокремлюється пізніше. У всіх безхребетних тварин, за винятком голкошкірих, вона утворюється з двох або декількох вихідних клітин - телобластів (телобластичний спосіб утворення мезодерми). У голкошкірих і всіх хордових, крім вищих хребетних, мезодерма виділяється з первинної ентодерми (ентероцельний спосіб). У плазунів, птахів і ссавців в процесі гаструляції мезодерма мігрує із епібласту (первинної ектодерми).
Нейруляція та утворення сомітів
Частини зародкових листків, які утворились внаслідок гаструляції, впливаючи один на одного, індукують утворення нових структур. Прикладом подібного впливу служить первинна ембріональна індукція. Її результатом є розвиток з дорзальної ектодерми нервової системи. Процес закладки нервової системи та осьових структур зародка, який у людини починається з 16 доби розвитку і в основному завершується до 22-23 доби, одержав назву нейруляція. Майже одночасно із мезодерми формуються соміти і нефротом.
Процес нейруляції відбувається у декілька стадій (Рис. 47):
1) індукція нервової пластинки;
2) підняття країв нервової пластинки і утворення нервового жолобка;
3) поява нервових валиків;
4) формування нервового гребеня початок виселення з нього клітин;
5) злиття нервових валиків - утворення нервової трубки;
6) змикання ектодерми над нервовою трубкою.
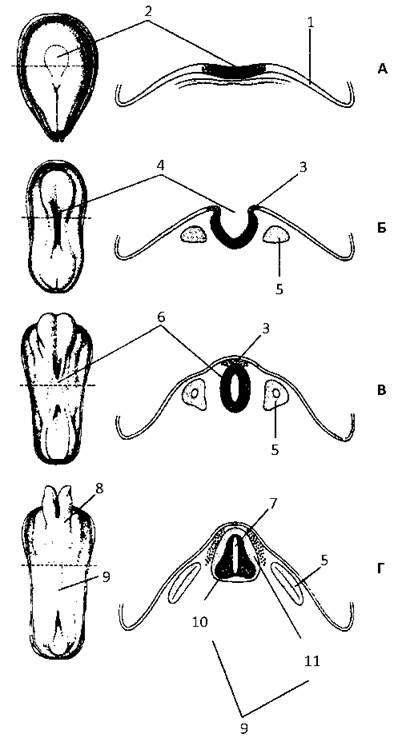
Рис. 47. Розвиток нервової системи:
А - нервова пластинка - 19 доба, Б - нервовий жолобок - 20 доба, В - нервова трубка - 22 доба, Г - зачаток ЦНС - 23-24 доба.
Зліва - дорзальна поверхня зародка, справа - дорзальна частина зародка на поперечному розтині, позначеному пунктиром зліва: 1 - ектодерма, 2 - нервова пластинка, 3 - нервовий гребінь, 4 - нервовий жолобок, 5 - соміт, 6 - нервова трубка, 7 - центральний канал, 8 - головний мозок, 9 - спинний мозок, 10 - сіра речовина, 11 — біла речовина.
Індуктивний вплив стосовно утворення нервової пластинки із дорзальної ектодерми здійснює хордомезодерма, яка у даному випадку виступає як організатор. В процесі первинної ембріональної індукції визначається подальша доля клітин, які дають початок нервовій системі. Природа індуктора та механізм індуктивної взаємодії між хордомезодермою і дорзальною ектодермою залишається поки нез’ясованим. Вважають, що клітини хордомезодерми виробляють хімічний фактор - індуктор. Цю гіпотетичну речовину називають нейралізуючим фактором.
З іншого боку, одержані дані про роль саме ектодермальних клітин в утворенні нервової системи. На ранніх ембріонах шпорцевої жаби показано, що ще до контакту з хордомезодермою ектодермальні клітини вже схильні до диференціювання у нервову тканину. Виявлено також, що в утворенні та подальшому диференціюванні нервової пластинки важливу роль відіграють інформаційні взаємодії між її клітинами через щілиноподібні контакти. Так, обробка зародків антитілами до білків нексусів запобігає утворенню нервової пластинки.
Первинна ембріональна індукція відбувається за краніокаудальним градієнтом, т.т. у напрямку від головного кінця зародка до хвостового. Під час нейруляції змінюється форма клітин дорзальної ектодерми - відбувається їхнє видовження у дорзовентральному напрямку, при цьому аналогічно орієнтуються і мікротрубочки їхньої цитоплазми. В апікальній частині клітини нервової пластинки з’єднані за допомогою щільних контактів, а у базальній - щілинних.
Невдовзі після утворення краї нервової пластинки припіднімаються і формують валики, між якими розташований нервовий жолобок. Пізніше краї нервових валиків змикаються по серединній лінії і утворюється замкнена нервова трубка.
Замикання нервової трубки відбувається не одночасно по довжині зародка. Нервова трубка замикається перш за все на рівні третьої пари сомітів, т.т. стовбуру майбутнього головного мозку (22 доба), а потім в інших відділах. В ембріональній нервовій системі є ще один регіон, де достатньо рано відбувається змикання нервової трубки - це рівень формування зачатків очей. Краніальна та каудальна ділянки нервової трубки ще деякий час залишаються незамкненими, їх називають відповідно переднім і заднім нейропорами. Передній нейропор закривається на 23-26 добу розвитку, а задній - на 26-30.
До 6-го тижня розвитку закінчується процес реорганізації каудальної частини нервової трубки, а до 8-го тижня утворюється кінській хвіст спинного мозку, що знаменує повне завершення нейруляції.
Не весь матеріал нервової пластинки входить до складу нервової трубки. Після змикання валиків і утворення нервової трубки частина ектодерми, розташована між нейральною і шкірною ектодермою, утворює нові структури - нервовий гребінь і нейрогенні плакоди.
Із матеріалу нервового гребеня в подальшому розвиваються чутливі нейрони спинномозкових вузлів і деяких гангліїв черепних нервів, нейрони автономної нервової системи, шваннівські клітини, меланоцити, клітини дифузної ендокринної системи (APUD-система), хрящі, кістки, м’язи і сполучна тканина обличчя, одонтобласти та ін.
Нейрогенні плакоди - це потовщення ектодерми, розташовані по обидва боки від нервової трубки у краніальному відділі зародка. Похідними нейрогенних плакод є нюхові нейрони, нейрони вестибулярного і слухового гангліїв, а також чутливі нейрони колінчастого, каменистого, вузлуватого і трійчастого гангліїв черепних нервів.
Практично одночасно з нейруляцією відбувається ще один досить важливий процес - диференціювання мезодерми і утворення осьового комплексу зачатків органів.
Із клітин первинної зародкової мезодерми формуються передсомітна мезодерма, з якої виникають соміти - симетричні парні структури по боках від хорди і нервової трубки, а також ще два великих зачатка - нефротом, або проміжна мезодерма, і латеральна мезодерма.
У безпосередній близькості від нервової трубки і хорди мезодермальні клітини утворюють скупчення - концентричні шари клітин метамерної організації у вигляді потенційних сомітів, або сомітомерів - передсомітну мезодерму. Вона виникає дуже рано - ще під час гаструляції. Сомітомери передсомітної мезодерми визначають сегментацію хорди, нервової трубки, проміжної мезодерми і латеральної мезодерми.
Клітини передсомітної мезодерми активно проліферують, і після їхньої міграції і агрегації утворюється дорзальна мезодерма - соміти (Рис. 48). Утворення сомітів відбувається за краніокаудальним градієнтом. Нова пара сомітів утворюється позаду останньої вже сформованої через певний проміжок часу. У зародка людини цей інтервал в середньому становить 6,6 години. В центрі соміта утворюється порожнина, а клітини, що її оточують, з’єднуються щільними контактами. В кожному соміті диференціюються ділянки склеротому, дерматому і міототому, які служать джерелами для розвитку різних структур зародка.
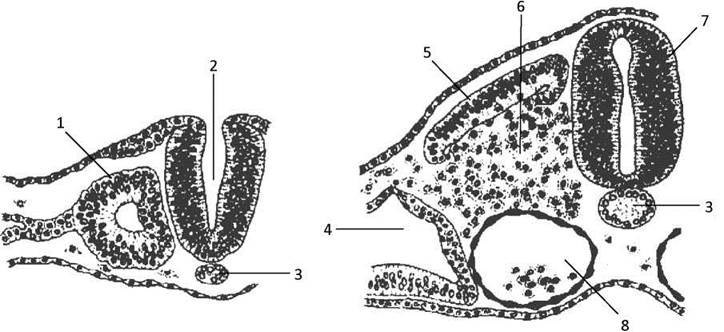
Рис. 48. Диференціювання мезодерми:
1 - соміт, 2 - нервовий жолобок, 3 - хорда, 4 - целом, 5 - дермоміотом, 6 - склеротом, 7 - нервова трубка, 8 - дорзальна аорта.
Склеротом складає вентромедіальну ділянку соміта. Його клітини під впливом матеріалу хорди і нервової трубки інтенсивно проліферують і виселяються із соміта, оточуючи хорду і вентральну частину нервової трубки. Клітини склеротому диференціюються у скелетогенні і утворюють хребці, ребра і лопатки.
Внутрішній шар решти дорзолатеральної частини соміта - міотом - утворює в подальшому скелетні м’язи, а зовнішній - дерматом - служить зачатком сполучнотканинного шару шкіри - дерми.
Латеральніше соміта розташовується ділянка проміжної мезодерми - нефротом (Рис. 49). Клітини нефротому служать зачатком сечостатевої системи.
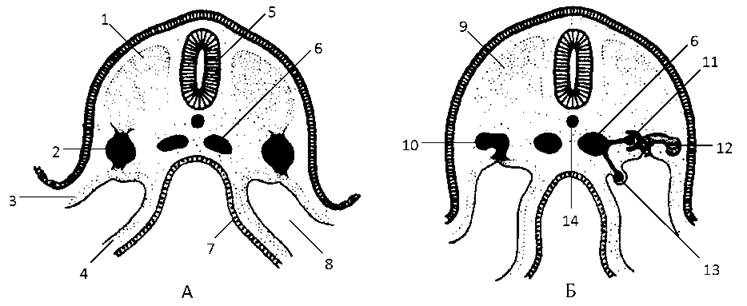
Рис. 49. Диференціювання проміжної мезодерми і нефротому:
А - 21-а доба, Б - 25-а доба розвитку зародка людини.
1 - дорзальна мезодерма, 2 - проміжна мезодерма, 3 - соматична мезодерма, 4 - спланхнічна мезодерма, 5 - нервова трубка, 6 - дорзальна аорта, 7 - ентодерма, 8 - целом, 9 - соміт, 10 - нефротом, 11 - внутрішній капілярний клубочок, 12 - каналець, 13 - зовнішній капілярний клубочок, 14 - хорда.
Крайову позицію у мезодермальній закладці займає пластинка латеральної мезодерми. Вона розщеплена на два листка: дорзальний і вентральний. Клітини дорзального листка (вживається також термін - парієтальний, т.т. пристінковий) - соматична мезодерма - в подальшому утворюють серозні оболонки (парієтальні плевра і очеревина).
Вентральний (вісцеральний - органний) листок - спланхнічна (гр. splanchna - нутрощі) мезодерма - дає початок багатьом структурам зародка і організму в цілому. Із неї утворюються: серце, кора надниркових залоз, строма гонад, сполучна і гладком’язова тканини внутрішніх органів (в т.ч. серозні оболонки) і кровоносних судин.