Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Клетки и молекулы за работой
Движение в немышечных клетках
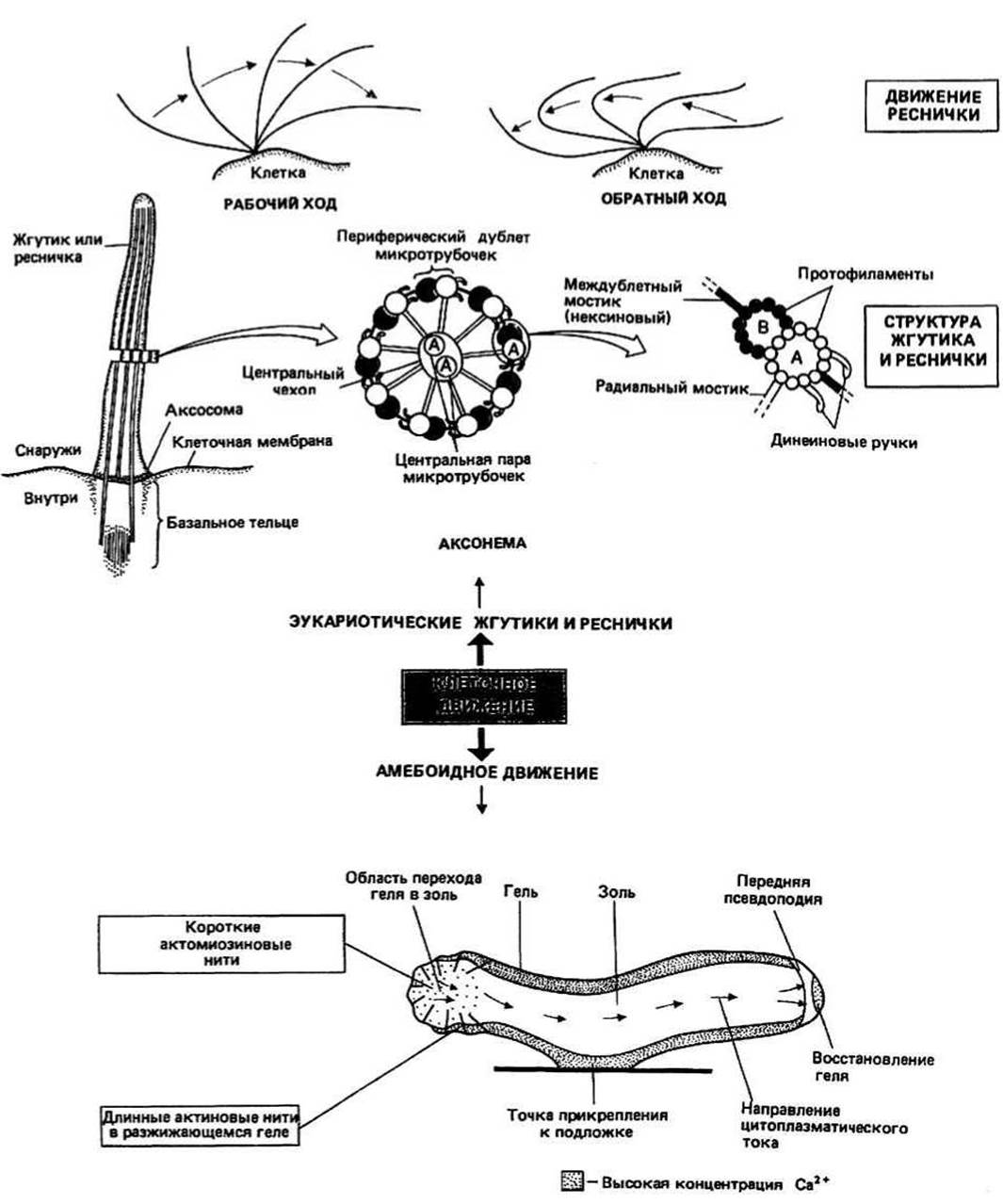
Рис. 39.1.
КЛЕТОЧНАЯ ПОДВИЖНОСТЬ - явление, свойственное как большинству одноклеточных эукариотических организмов, так и многим типам клеток многоклеточных. Хотя простейшие, клетки растений и клетки животных значительно различаются по фенотипу, механизмы, управляющие их подвижностью, обнаруживают замечательное сходство. Тот же клеточный аппарат, который контролирует движение клетки в целом, способен управлять и движением органелл внутри клетки. Все типы клеточной подвижности можно классифицировать в зависимости от того, обеспечиваются ли они микротрубочками или микро - филаментами.
MИКРОТРУБОЧКИ — длинные трубчатые структуры. Каждая микротрубочка имеет постоянный диаметр и состоит из молекул глобулярного белка тубулина, являющегося димером с мол. массой 110 000—120 000. Существует два типа мономеров тубулина, ос и р, и димер является обычно гетеродимером — ар. Микротрубочка представляет собой полый цилиндр с внешним диаметром около 24 нм и внутренним диаметром 15 нм. Длина микротрубочек варьирует от нескольких микрометров в цитоплазме большинства клеток и нескольких десятков микрометров в жгутиках и ресничках до нескольких миллиметров в аксонах нервов (гл. 36). Стенка цилиндра, имеющая толщину около 5 нм, состоит из 13 расположенных по кругу, соприкасающихся бок о бок протофиламентов, каждый из которых является полимером тубулинового гетеродимера. Вид такой микротрубочки сбоку показан на рис. 39.2. В эукариотических клетках микротрубочки могут входить в состав характерной структуры, аксонемы жгутиков и ресничек, отличной от структур, встречающихся в прокариотических клетках (гл. 2). Поскольку жгутики и реснички есть как у одноклеточных организмов (например, у простейших), так и у многоклеточных (например, у человека) и играют существенную роль в клеточном движении, строение аксонемы будет рассмотрено в деталях (см. ниже). Находясь в цитоплазме эукариотических клеток, микротрубочки играют ключевую роль в таких процессах, как образование митотического веретена и эндоцитоз (гл. 3), а в жгутиках и ресничках они участвуют в генерации движения клеток.
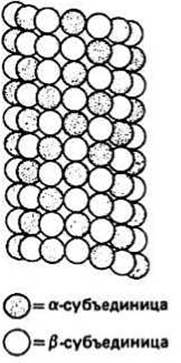
Рис. 39.2.
Белки, ассоциированные с микротрубочками, — неотъемлемые компоненты системы микротрубочек; они играют существенную роль в регуляции таких процессов, как сборка и разборка микротрубочек и образование поперечных сшивок, связывающих микротрубочки между собой, а также с другими компонентами клетки.
Аксонема — это состоящая главными образом из микротрубочек структура, которая служит основным двигательным элементом жгутиков и ресничек. Она окружена мембраной — непосредственным продолжением плазматической мембраны клетки. Ресничками называются короткие (1—5 мкм) подвижные отростки клетки, присутствующие на ее поверхности в большом числе. Если же отростков лишь несколько и они длинные (до 1—2 мм), то их называют жгутиками. На поперечном срезе аксонемы видна центральная пара трубочек, окруженная кольцом из девяти дублетов трубочек. Аксонемные трубочки являются по своему строению либо классическими микротрубочками со стенкой, построенной из 13 протофиламентов (трубочки типа А), либо неполными трубочками из 10 протофиламентов, образующих незамкнутый - серповидный на поперечном срезе — цилиндр (трубочки типа В). В непосредственной близости от двух центральных трубочек, которые принадлежат к типу А, располагается окружающий их белковый чехол. Несколько далее находятся девять дублетов трубочек, каждый из которых состоит из трубочки типа А и присоединенной к ней трубочки типа В. В аксонеме имеются два типа постоянных мостиков: междублетные мостики, которые построены из белка нексина и соединяют А-трубочки дублетов с В-трубочками соседних дублетов, и радиальные мостики, которые состоят из не имеющего еще названия белка и связывают каждый дублет с центральным чехлом. На А-трубочках дублетов расположен с некоторым периодом белок динеин; в определенных условиях он образует междублетные мостики, которые и генерируют силу, приводящую жгутики и реснички в движение. Хотя по характеру движения реснички и жгутики немного различаются - у первых оно похоже на удар хлыста, а у вторых является волнообразным, — его молекулярная основа и в том и в другом случае одна и та же.
Модель скользящих нитей, предложенная для объяснения движения ресничек, имеет много общего с одноименной моделью, описывающей мышечное сокращение (гл. 37). В присутствии АТР белок динеин, образующий «ручки» на А-трубочке каждого дублета, связывается с В-трубочкой соседнего дублета. Этот белок является Са2+- и Mg+-зависимой АТРазой высокой мол. массы (300 000—400 000) и может находиться по меньшей мере в двух конформационных состояниях. При связывании с соседней В-трубочкой он находится в одном состоянии, а во время последующего гидролиза АТР переходит в другое. Этот переход, сопровождающийся движением динеиновых ручек в плоскости длинной оси трубочек, приводит к смещению одних трубочек относительно других. В результате образования и диссоциации множества мостиков трубочки скользят, или, точнее, «ползут», вдоль соседних трубочек. Если скольжение будет происходить преимущественно на одной стороне аксонемы, ресничка изогнется. В верхней части рис. 39.1 показан ряд последовательных положений реснички, изгибающейся в процессе биения. Бьющими ресничками клетка отталкивается от жидкой среды и таким образом перемещается в ней.
МИКРОФИЛАМЕНТЫ — нитевидные органеллы диаметром около 5 нм — встречаются в большинстве эукариотических клеток; они являются полимером глобулярного белка актина (мол. масса 43 000). Микрофиламенты располагаются в цитоплазме беспорядочно либо образуют фибриллярные пучки, например в выступающих вперед отростках движущихся клеток. У многих клеток млекопитающих пучки микрофиламентов, называемые иногда «волокнами натяжения», формируют во время интерфазы (гл. 29) сеть, простирающуюся по всему внутриклеточному пространству. Во время митоза многочисленные микрофиламенты обнаруживаются в зоне, где происходит разделение клетки на дочерние; микрофиламенты образуют там так называемое «сократительное» кольцо. Немышечные клетки содержат также белок миозин, и в основе их движения может, по-видимому, лежать тот же тип взаимодействия актина и миозина, что и в основе мышечного сокращения. Одна из г ипотез о механизме движения амебы, а именно предложенная Тейлором модель разжижения—сокращения, рассматривается ниже.
Модель разжижения - сокращения, описывающая механизм амебоидного движения, постулирует, что цитоплазма амебы может находиться в двух состояниях — геля и золя. Гелеобразная цитоплазма локализуется лишь в областях, непосредственно прилегающих к плазматической мембране, и содержит актиновые нити, сшитые друг с другом актинсвязывающими белками одного или нескольких типов (например, ос-актинином или филамином). Из-за наличия поперечных сшивок актиновые нити геля, хотя и контактируют с миозином, не могут участвовать в сокращении. В задней части амебы имеется область высокой концентрации Са2+, вызывающей отсоединение актинсвязывающих белков от актиновых нитей; это область перехода геля в золь. После отделения сшивающих белков другие белки (например, виллин и гельзолин) дестабилизируют длинные актиновые нити, и в результате образуются короткие нити, способные взаимодействовать с миозином. Взаимодействие с миозином приводит к сокращению, протекающему согласно классической модели скользящих нитей (гл. 37). Результатом сокращения в задней части амебы, там, где гель переходит в золь, является повышение гидравлического давления, что заставляет цитоплазму амебы перемещаться к переднему краю, в выступающую вперед псевдоподию. Как только растворимый актин и актинсвязывающие белки достигают передней части клетки, они вновь образуют гель, поддерживая таким образом постоянство соотношения между гелем и золем в амебе. Аналогичные схемы, предполагающие перемещение цитоплазмы — «цитоплазматический ток» — за счет возникающих при сокращении «толкающих» сил, предложены для объяснения движения у многих клеток высших организмов.