Биологические мембраны - А. Н. Огурцов 2012
Структура и функции биомембран
Структура липидных мембран
Жидкокристаллическое состояние биомембран
Принципиальное отличие твёрдого тела от жидкости заключается в наличии или отсутствии дальнего порядка и в том, какое время молекулы вещества проводят вблизи положений равновесия между перескоками. Время оседлой жизни молекулы в жидкости много меньше, чем в твёрдом теле.
Липидные бислойные мембраны при физиологических условиях - жидкие, время оседлой жизни фосфолипидных молекул в мембране мало: τ ≈10-7 - 10-8 с. Вместе с тем, молекулы в мембране размещены не беспорядочно, в их расположении наблюдается дальний порядок. Фосфолипидные молекулы находятся в двойном слое, а их гидрофобные хвосты приблизительно параллельны друг другу. Есть порядок и в ориентации полярных гидрофильных голов.
Физическое состояние, при котором есть дальний порядок во взаимной ориентации и расположении молекул, но агрегатное состояние жидкое, называется жидкокристаллическим состоянием.
Жидкие кристаллы могут образовываться не во всех веществах, а в веществах из "длинных молекул" (поперечные размеры которых меньше продольных) (рисунок 23).
Могут быть различные жидкокристаллические структуры:
✵ нематическая (нитевидная), когда длинные молекулы ориентированы параллельно друг другу (рисунок 23(6));
✵ смектическая (мылообразная) - молекулы параллельны друг другу и располагаются слоями (рисунок 23(b));
✵ холестерическая - молекулы располагаются параллельно друг другу в одной плоскости, но в разных плоскостях ориентации молекул разные (повернуты на некоторый угол в одной плоскости относительно другой) (рисунок 23 (г)).
Бислойная липидная фаза биологических мембран соответствует смектическому (рисунок 23(b)) жидкокристаллическому состоянию.
Жидкокристаллические структуры чувствительны к изменению температуры, давления, химического состава, к наличию электрического поля. Это определяет динамичность липидных бислойных мембран - изменение их структуры при различных, даже небольших изменениях внешних условий или химического состава.
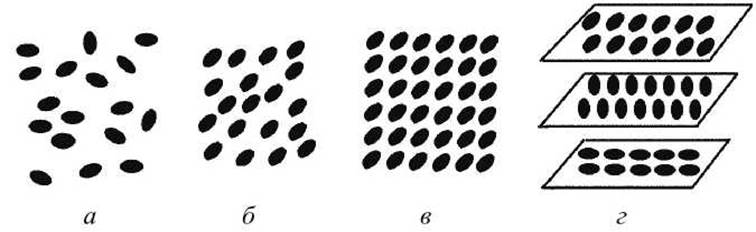
Рисунок 23 - Возможные схемы расположения молекул в жидких кристаллах: а - в аморфной фазе, б - в нематическом состоянии, в - в смектическом состоянии, г - в холестерическом состоянии
Степень вязкости жидкокристаллической фазы мембраны и возможность её перехода в гелеобразное состояние зависят от липидного состава мембраны, структуры гидрофобных хвостов фосфолипидов и от температуры.
Гидрофобный эффект и ван-дер-ваальсовые взаимодействия вызывают агрегацию неполярных хвостов фосфолипидов. Причём для длинных хвостов с насыщенными С-С связями наблюдается максимальная агрегация, приводящая к образованию гелеобразных мембран. А фосфолипиды с более короткими жирнокислотными хвостами, у которых гораздо меньшая площадь взаимодействий с соседями, образуют более жидкие бислои.
Аналогично, и жёсткие изгибы жирнокислотных хвостов (кинки) вследствие наличия ненасыщенных С=С связей приводят к ослаблению ван-дер-ваальсовых связей с соседями (по сравнению с насыщенными аналогами) и, следовательно, к "разжижению" бислоя.
Так, например, добавление ненасыщенной связи в 18-углеродный хвост стеариновой кислоты, температура плавления которой +69,6°С (рисунок 24(a)), приводит к образованию олеиновой кислоты, температура плавления которой +13,4°С (рисунок 24(6)). 18-Углеродная линолевая кислота, у которой две ненасыщенных связи в хвосте, имеет температуру плавления -5°С (рисунок 24(b)), а у линоленовой (рисунок 24(г)) и её изомера у-линоленовой (рисунок 24(д)) кислоты с тремя кинками в хвосте температура плавления равна -11 °С.
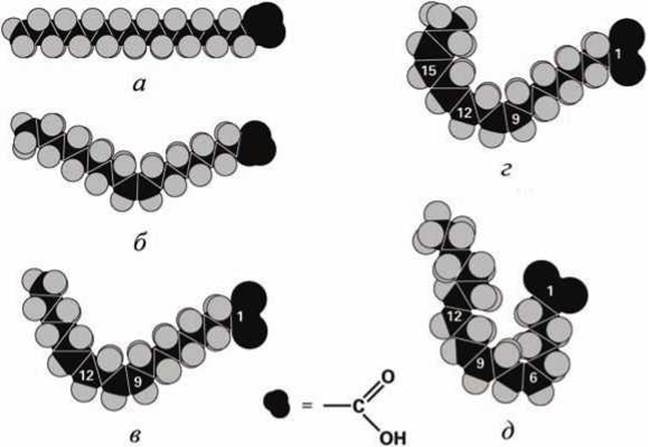
Рисунок 24 - 18-Углеродные жирные кислоты: а - стеариновая, б - олеиновая, в - линолевая, г - линоленовая, д - у-линоленовая
Нагрев высокоупорядоченного гелеобразного бислоя приводит к увеличению амплитуды движений в жирнокислотных хвостах липидов, что приводит к переходу в более жидкое, разупорядоченное состояние - фазовый переход гель —> жидкий кристалл (рисунок 25).
При физиологических температурах внутренний гидрофобный слой природных мембран, как правило, имеет низкую вязкость и является жидкостью.
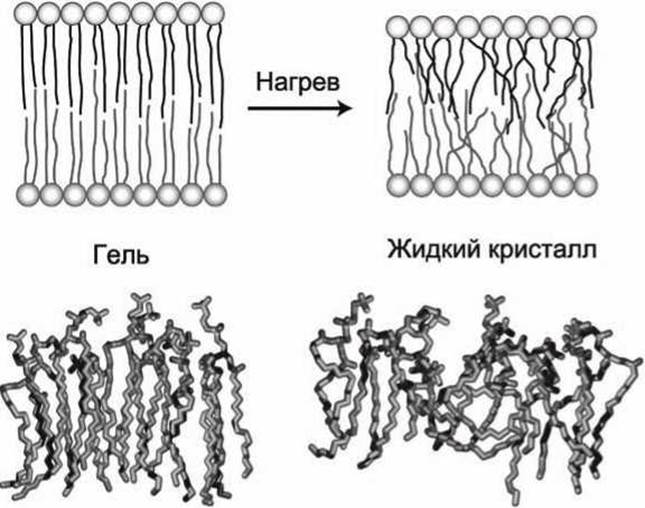
Рисунок 25 - Фазовый переход гель-жидкий кристалл при нагреве мембраны
В гель-состоянии молекулы расположены ещё более упорядочено, чем в жидкокристаллическом. Все гидрофобные углеводородные хвосты фосфолипидных молекул в гель-фазе полностью вытянуты строго параллельно друг другу (имеют полностью транс-конформацию).
В жидком кристалле за счёт теплового движения возможны транс-гош-переходы, хвосты молекул изгибаются, их параллельность друг другу в отдельных местах нарушается, причём особенно сильно в середине мембраны. Это приводит к тому, что толщина мембраны в гель-фазе больше, чем в жидком кристалле.
При переходе из твёрдого в жидкокристаллическое состояние объём мембраны несколько увеличивается, поскольку значительно увеличивается площадь мембраны, приходящаяся на одну молекулу (от 0,48 нм2 до 0,58 нм2). Для нормального функционирования мембрана должна быть в жидкокристаллическом состоянии.
Поэтому в живых системах при продолжительном понижении температуры окружающей среды наблюдается адаптационное изменение химического состава мембран, обеспечивающее понижение температуры фазового перехода. Температура фазового перехода понижается при увеличении числа ненасыщенных связей в жирно-кислотных хвостах. В хвосте молекулы может быть до четырёх ненасыщенных связей. В зависимости от химического состава липидных мембран температура фазового перехода гель—жидкий кристалл может меняться от - 20 °С (для мембран из ненасыщенных липидов) до +80°С (для насыщенных липидов).
У микроорганизмов, растительных и животных клеток наблюдается увеличение числа ненасыщенных липидов в мембране при понижении температуры обитания.
Характерный пример приспособления клеточных мембран к температурным условиям — изменение температуры фазового перехода (за счёт изменения химического состава мембранных липидов) ноги полярного оленя. Температура вдоль ноги полярного оленя от копыта до туловища может зимой меняться от -20 °С до +30°С. Клеточные мембраны у дистальной части ноги оленя содержат больше ненасыщенных фосфолипидов.
Предполагается, что первичный механизм криоповреждений (повреждений при охлаждениях) биологических мембран связан с фазовым переходом в гель-состояние. Поэтому биологические мембраны содержат большое количество холестерола, уменьшающего изменения в мембране, сопровождающие фазовый переход.
При фазовых переходах из гель- в жидкокристаллическое состояние и обратно в липидном бислое могут образовываться сквозные каналы, радиусом ~2 нм, по которым через мембрану могут переноситься ионы и низкомолекулярные вещества. Вследствие этого при температуре фазового перехода может резко увеличиваться ионная проводимость мембраны. Увеличение ионной проводимости мембран может спасти клетку от криоповреждений за счёт увеличения выхода из клетки воды и солей, что препятствует кристаллизации воды внутри клетки.
У некоторых микроорганизмов биологические мембраны находятся при температурах, лишь немного превышающих температуру фазовых переходов липидов. Возможно, повышение ионной проводимости мембран при фазовом переходе позволяет поддерживать метаболический обмен некоторых микроорганизмов. Большой интерес представляет этот эффект для объяснения термо- и хеморецепции.
Перенос ионов через мембрану лежит в основе формирования биопотенциалов, изменение ионной проводимости обусловливает нервный импульс. Не исключено, что нервный импульс, свидетельствующий о понижении или повышении температуры (терморецепция), образуется за счёт изменения ионной проницаемости липидного бислоя при фазовом переходе мембранных липидов.
Некоторые виды хеморецепции также могут быть связаны с фазовым переходом мембранных липидов, поскольку фазовый переход может быть вызван не только изменением температуры, но и изменением химического состава окружающей среды. Например, при данной температуре фазовый переход из жидкокристаллического состояния в гель-состояние может быть вызван увеличением концентрации Са2+ в физиологическом диапазоне от 1 до 10 ммоль/л в водном растворе, окружающем мембрану.
Для управления вязкостью мембран клетки используют прежде всего холестерол. Холестерол не может самостоятельно образовать бислой. При физиологических концентрациях холестерол интеркалирует (внедряется) между фосфолипидами. Холестерол ограничивает случайное движение полярных головок фосфолипидов на наружных сторонах мембраны, а его влияние на движение гидрофобных хвостов фосфолипидов зависит от концентрации.
При обычных концентрациях холестерола взаимодействие стероидных циклов с длинными гидрофобными хвостами фосфолипидов затрудняет их свободное движение, что увеличивает вязкость мембран. При снижении концентрации холестерола стероидные кольца отсоединяются от липидных хвостов и даже расталкивают их, способствуя диспергированию липидной фазы, что приводит к снижению вязкости биомембран.