ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 25. Гиперчувствительность - тип III
ПЕРСИСТЕНЦИЯ КОМПЛЕКСОВ
В норме иммунные комплексы уничтожаются мононуклеарными фагоцитами
Иммунные комплексы после активации комплемента опсонизируются С3b и уничтожаются мононуклеарными фагоцитами, преимущественно в печени и селезенке. Элиминация происходит при участии С3b-рецепторов (CR1). У приматов CR1 присутствует в основном на эритроцитах. (У других животных эритроциты лишены CR1 и поэтому опсонизированные комплексы связываются с CR1 тромбоцитов.) На каждом эритроците присутствует примерно 700 рецепторов и их эффективность повышается за счет кластеризации, что создает возможность высокоавидиого связывания крупных комплексов. Как показывают эксперименты на животных, лишенных комплемента, рецепторы CR1 легко связывают лишь те иммунные комплексы, которые фиксировали комплемент (рис. 25.11).
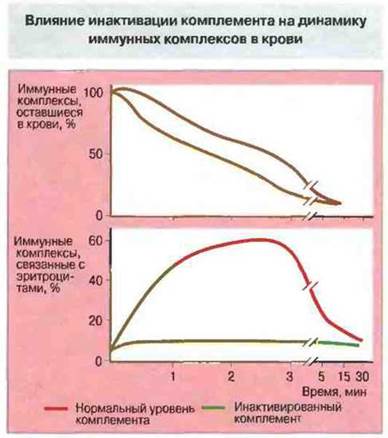
Рис. 25.11. Обезьянам одномоментно впрыскивали в кровь иммунные комплексы. У животных с нормальной системой комплемента они быстро связывались CR1 на поверхности эритроцитов. В то же время у животных, которым предварительно вводили яд кобры для инактивации комплемента, эритроциты практически не связывали иммунные комплексы. Парадоксально, но это несколько ускоряло исчезновение комплексов из крови; они, однако, не элиминировались клетками селезенки, а откладывались в тканях. (По Waxman et at., 1984.)
В норме у приматов эритроциты выполняют функцию буфера, связывая фиксировавшие комплемент комплексы и эффективно удаляя их из плазмы. В мелких сосудах ток крови ламинарный, при этом эритроциты локализованы в центре сосуда и со всех сторон окружены плазмой. Таким образом, со стенками сосуда контактирует только плазма (рис. 25.12). Эритроциты соприкасаются с внутренней оболочкой сосудов лишь в синусоидах печени и селезенки, где ток крови становится турбулентным.
Комплексы переносятся в печень и селезенку, где их извлекают из крови тканевые макрофаги (рис. 25.12). При этом удаляется и большая часть CRI: поэтому в случае постоянного образования иммунных комплексов количество активных рецепторов на эритроцитах постепенно уменьшается, что снижает эффективность транспорта комплексов. У больных СКВ, например, количество рецепторов может быть снижено вдвое.
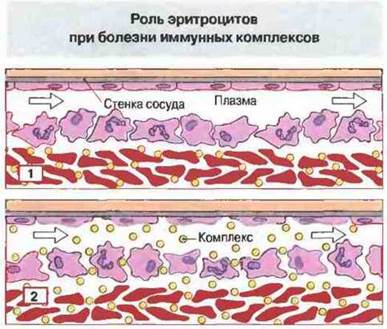
Рис. 25.12. В сосудах кровь течет ламинарным потоком и в центре его находятся эритроциты, окруженные лейкоцитами и затем еще как бы чехлом из плазмы, которая контактирует с сосудистой стенкой. Иммунные комплексы, связанные с эритроцитами через CR1 (1), удерживаются в отдалении от стенок сосуда. Недостаточность комплемента препятствует связыванию комплексов с эритроцитами и создает возможность контакта и связывания комплексов со стенками сосуда (2).
Комплексы могут также отделяться от эритроцитов в крови благодаря ферментативному действию фактора I, который расщепляет С3b, оставляя присоединенным к CR1 на клеточной мембране небольшой его фрагмент (C3dg). Эти растворенные комплексы удаляются затем фагоцитами, несущими рецепторы для Fc-фрагмента IgG (рис. 25.13).
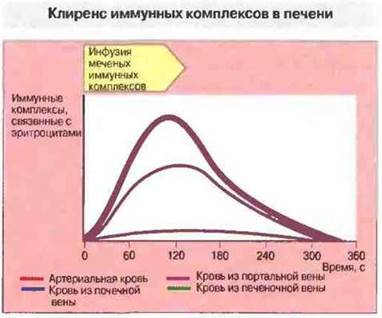
Рис. 25.13. Комплексы 125I-БСА/анти-БСА вводили обезьянам в течение 120 с. Содержание связанных с эритроцитами комплексов определяли радиометрически в пробах крови из почечной, портальной и печеночной вен. Содержание комплексов в почечной и портальной венах не отличалось от их содержания в артериальной крови. Однако в крови печеночных вен комплексы практически отсутствовали. Это свидетельствует об удалении связанных с эритроцитами комплексов за время одного пассажа через печень. (Пo Cornacoff etal., 1983.)
Солюбилизация иммунных комплексов комплементом. Еще в 1930-х гг. Гейдельбергер (Hcidclbcrger) обнаружил, что комплемент замедляет преципитацию иммунных комплексов. (Эти данные были потом надолго забыты.) Удержание иммунных комплексов в растворимом состоянии обеспечивается действием классического пути активации комплемента. Белки комплемента снижают количество антигенных эпитопов (т. е. валентность антигена), с которыми смогут связываться антитела, внедряясь в структуру комплекса. Это приводит к уменьшению размеров комплексов и делает их растворимыми. У приматов такие содержащие комплемент комплексы легко связываются С3b-рецепторами (CR1) на поверхности эритроцитов.
Комплемент способен быстро ресолюбилизировать осажденные комплексы благодаря альтернативному пути активации (рис. 25.14). Солюбилизация происходит, по-видимому, в результате встраивания в комплекс фрагментов комплемента С3b и C3d.
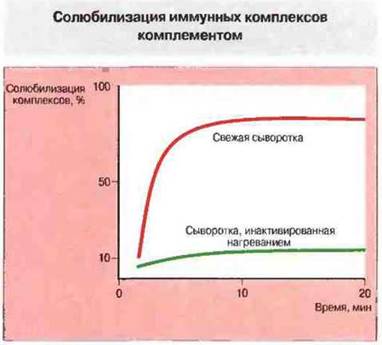
Рис. 25.14. Комплемент способен солюбилизировать осаждаемые комплексы in vitro. Добавление свежей сыворотки, содержащей активный комплемент, к нерастворимым комплексам (при 37 °С) вызывает уже через 15 мин их солюбилизацию. Часть комплексов остается резистентной к солюбилизации. Прогретая сыворотка (56 °С в течение 30 мин) лишена активного комплемента и не способна солюбилизировать комплексы. Включение компонентов комплемента С3b и C3d в комплексы приводит к солюбилизации в результате разрыва связей между антигеном и антителами. Искусственно образованные комплексы с ковалентными связями не поддаются солюбилизирующему действию комплемента.
Возможно, у здоровых людей непрерывно происходит отложение комплексов, но они столь же непрерывно удаляются путем солюбилизации. Если это так, то при сниженном содержании комплемента процесс должен нарушаться и комплексы могут накапливаться в тканях. Действительно, у лиц, страдающих системной болезнью иммунных комплексов, солюбилизирующий эффект сыворотки снижен по сравнению с нормой, хотя неизвестно, является это причиной или следствием заболевания.
При недостаточности комплемента нарушается клиренс комплексов. У лиц с низким уровнем компонентов классического пути активации комплемента иммунные комплексы плохо связываются эритроцитами. Недостаточность комплемента может быть следствием его исчерпания, вызванного болезнью иммунных комплексов, или врожденным состоянием, как это имеет место в случае дефицита С2. Можно было бы ожидать, что в таких случаях иммунные комплексы постоянно будут находиться в крови, но на самом деле происходит обратное: комплексы быстро исчезают из крови. Такие не связанные с эритроцитами комплексы активно поглощаются печенью (но не селезенкой), а затем высвобождаются вновь и откладываются в таких тканях, как кожа, почки и мышцы, вызывая в них воспалительные реакции (рис. 25.15).
Вливание свежей плазмы, содержащей комплемент, восстанавливает нормальный клиренс иммунных комплексов, что свидетельствует о роли комплемента в этом процессе. Отсутствие поглощения комплексов в селезенке не только приводит к болезни иммунных комплексов, но может иметь важное значение и для развития соответствующих иммунных реакций, поскольку селезенке принадлежит центральная роль в процессинге антигенов и индукции иммунного ответа (см. гл. 3).
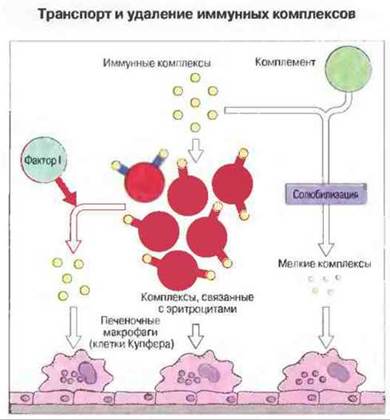
Рис. 25.15. У приматов комплексы связываются с CR1 на эритроцитах и поступают в печень, где их поглощают тканевые макрофаги. Комплексы, отделившиеся от эритроцитов под действием фактора I, связываются клетками (в том числе и макрофагами), обладающими рецепторами к Fc и комплементу. Солюбилизация крупных комплексов комплементом превращает их в меньшие по размерам растворимые частицы, которые могут непосредственно поглощаться тканевыми макрофагами.
Отложение иммунных комплексов зависит от их размеров. Крупные иммунные комплексы, как правило, уже за несколько минут извлекаются из крови в печени, тогда как комплексы меньших размеров остаются в крови более длительное время (рис. 25.16). Это объясняется тем, что крупные комплексы эффективнее фиксируют комплемент и за счет этого лучше связываются с эритроцитами. Они и медленнее отделяются от эритроцитов под действием фактора I. Поэтому вес, что влияет на размер комплексов, должно изменять их клиренс. Как предполагается, генетический дефект, способствующий образованию низкоаффинных антител, вполне может обусловливать формирование мелких комплексов и тем самым развитие болезни иммунных комплексов. Повышение (созревание) аффинности зависит от эффективного соматического мутирования и селекции В-клеток в центрах размножения после связывания антигена. Этот процесс протекает гораздо эффективнее в тех случаях, когда В клетки стимулируются антигеном или иммунными комплексами, покрытыми комплементом. Недостаточность комплемента увеличивает предрасположенность к болезни иммунных комплексов, и одной из причин этого, как показывают недавно полученные данные, является замедленная скорость поступления связанного в комплексы антигена в центры размножения, препятствующая созреванию аффинности. Антитела к аутоантигенам могут обладать низкой аффинностью и распознавать лишь небольшое количество эпитопов. Этот дефект приводит к образованию мелких комплексов и возрастанию периода их клиренса, поскольку ограничивает формирование крупных перекрестно-связанных структур.
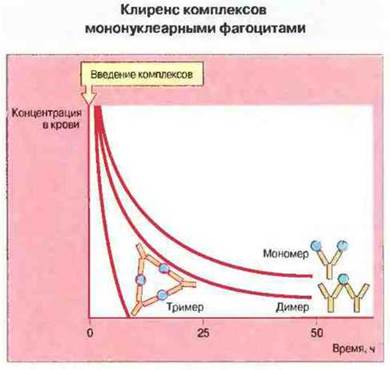
Рис. 25.16. Крупные иммунные комплексы удаляются из крови быстрее за счет того, что клетки ретикулоэндотелиальной системы, обладающие Fc-рецепторами, с большей авидностъю связывают Fc-фрагменты IgG. Кроме того, крупные комплексы лучше фиксируют комплемент, чем мелкие.
Скорость клиренса иммунных комплексов зависит от класса иммуноглобулинов. Класс иммуноглобулинов существенно влияет на клиренс комплексов. IgG-комплексы связываются с эритроцитами и удаляются из крови постепенно, тогда как IgA-комилсксы плохо связываются с эритроцитами, но исчезают из крови быстрее, в большей степени откладываясь в почках, легких и мозге.
Дефекты фагоцитов могут быть причиной персистенции комплексов. Опсонизированные иммунные комплексы в норме элиминируются мононуклеарными фагоцитами, главным образом в печени и селезенке. Однако при большом количестве комплексов возможна перегрузка системы мононуклеарных фагоцитов, приводящая к увеличению содержания комплексов в крови и большему их отложению в почечных клубочках и других тканях. При болезни иммунных комплексов у человека наблюдаются дефекты мононуклеарных фагоцитов, по это вполне может быть следствием их перегрузки, а не первичным нарушением.
На клиренс комплексов влияет углеводный компонент антител. Как установлено, важную роль в удалении иммунных комплексов фагоцитарными клетками выполняют углеводные группы молекул иммуноглобулинов При болезнях иммунных комплексов, например при ревматоидном артрите, наблюдаются отклонения в составе этих углеводов, приводящие к обострению патологического процесса. И з-за отсутствия концевого остатка галактозы в олигосахарид ной цепи Fc-фрагмента иммуноглобулинов усиливается связывание ревматоидного фактора. Недавно обнаружено, что с антителами IgG, не содержащими этого остатка галактозы, взаимодействует маннан-связывающий белок и в результате происходит активация комплемента.