ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 5. Миграция клеток и воспаление
МЕХАНИЗМЫ КЛЕТОЧНОЙ МИГРАЦИИ
Выход лейкоцитов из сосудистого русла через эндотелий происходит в несколько этапов (первые три из них показаны на рис. 5.10).
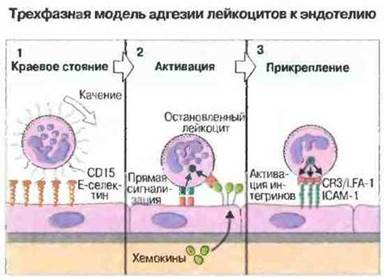
Рис. 5.10. Три фазы адгезии лейкоцитов на примере нейтрофила (у лейкоцитов других типов и в иных ситуациях действу ют другие наборы молекул межклеточной адгезии). 1. Краевое стояние, связывание лейкоцитарного CD15 с Е-селектином приводит к замедлению и к постепенной остановке нейтрофила, когда он катится по эндотелию. 2. Активация, задержанный нейтрофил активируется, непосредственно взаимодействуя с компонентами поверхности клеток эндотелия или с хемокинами и другими хемотаксическими молекулами, присутствующими на эндотелии. 3. Прикрепление: активация мобилизует лейкоцитарные интегрины (CR3 и LFA-1) для связывания с ICAM-1, экспрессия которой на поверхности эндотелия индуцируется при активации.
✵ Краевое стояние: движение лейкоцитов по венуле замедляется и они катятся но эндотелию. Это обусловлено в основном взаимодействием селектинов с углеводными лигандами.
✵ Активация: остановленные лейкоциты подвергаются воздействию цитокинов, хемотаксических агентов, компонентов поверхности эндотелия и внеклеточною матрикса. Эти факторы способны активировать клетку и включить программу ее миграции.
✵ В результате прикрепления лейкоцитов и мобилизации интегринов повышается аффинность лейкоцитарных интегринов, которые взаимодействуют с эндотелиальными молекулами межклеточной адгезии и активируют клетку для миграции.
✵ Миграция: при участии нового набора молекул адгезии лейкоциты связываются с базальной мембраной эндотелия и проникают сквозь нее.
✵ Лизис: мигрирующие клетки выделяют ферменты, лизирующие коллаген и другие компоненты базальной мембраны эндотелия; это позволяет лейкоцитам проникнуть в ткань.
Молекулы межклеточной адгезии регулируют прилипание лейкоцитов и прохождение их через эндотелий
Для каждого типа перемещения клеток характерно участие специфического набора молекул адгезии и хемотаксических агентов.
Миграция лейкоцитов к очагу воспаления. Нейтрофилы появляются в очаге острого воспаления на его ранней стадии, и отчасти это обусловлено индукцией цитокинами экспрессии Е-селектина на поверхности эндотелия в этой области. Стимуляция клеток эндотелия in vitro такими цитокинами, как, например, фактор некроза опухолей а (ФНОα) или интерлейкин I (ИЛ-I) индуцирует депрессию Е-селектина спустя 4-12 ч. а через 24 ч она прекращается (рис. 5.11); аналогичным образом in vivo эта молекула межклеточной адгезии появляется на ранней стадии воспалительной реакции. Клетки, трансфицированные геном Е-селектина, экспрессируют его в значительном количестве, приобретая при этом выраженную способность связывать нейтрофилы. Все эти данные укалывают, что остановка нейтрофилов при участии Е-селектина - это первая необходимая стадия их миграции.
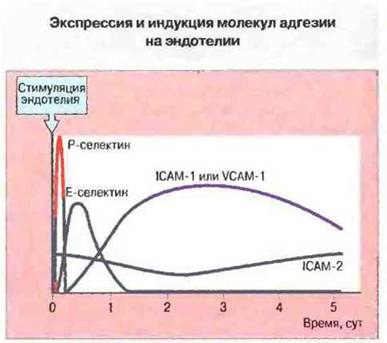
Рис. 5.11. Динамика экспрессии различных молекул межклеточной адгезии на клетках эндотелия пупочной вены человека после стимуляции ФНОα in vitro.
Важную роль в миграции нейтрофилов, лимфоцитов и моноцитов выполняют также экспрессируемые на лейкоцитах интегрины LFA-I и CR3, которые связываются с эндотелиальными молекулами межклеточной адгезии из суперсемейства иммуноглобулинов. Так, LFA-I связывается с ICAM-1 и ICAM-2 на эндотелии сосудов. В культуре клетки эндотелия конститутивно синтезируют ICAM-2; в связи с этим высказано предположение, что именно данный белок определяет фоновый уровень связывания лимфоцитов с эндотелием различных типов in vivo. Например, уровень экспрессии ICAM-2 на эндотелии мозговых сосудов в норме относительно низок и этому соответствует весьма незначительная трансэндотелиальная миграция лимфоцитов. Напротив, экспрессия ICAM-1, в норме низкая па поверхности эндотелия, может быть резко повышена цитокинами (ФНОα, ИЛ-I или ИФγ, в зависимости от вида животных). В условиях in vitroиндуцированная экспрессия IСАМ-1 наблюдается в период 8—96 ч после стимуляции (рис. 5.11), что соответствует более позднему прибытию в очаг воспаления in vivo лимфоцитов и моноцитов. Роль CR3 в привлечении фагоцитов показали опыты in vivo с использованием антител анти-СК3, которые, как было при этом установлено, подавляют миграцию данных клеток. У больных с дефицитом лейкоцитарной адгезии, подверженных в результате слабого накопления фагоцитов тяжелым инфекционным заболеваниям, отмечена недостаточность всех β2-интегринов (LFA-1, CR3, CR4). Следует отметить, что CR3 и LFA-1 связываются с разными участками ICAM-1.
Экспрессия VCAM-1, как и ICAM-I, индуцируется в области воспаления, причем in vitro индукция этих двух молекул происходит синхронно (рис. 5.11). (VCAM-1 связывается с интегрином α4β1, который экспрессируют лимфоциты некоторых субпопуляций, а также опосредует избирательную адгезию базофилов и эозинофилов, играя тем самым важную роль в аллергических реакциях.) Вместе с тем механизмы индукции Е-селектина. ICAM-1 и VCAM-1 у разных популяций лимфоцитов и клеток эндотелия на различных участках сосудистого русла тонко различаются. Это обеспечивает точную настройку миграции лейкоцитов сквозь эндотелий при воспалении и последовательное прибытие в очаг различных клеточных популяций. Прилипание лимфоцитов к эндотелию можно подавить антителами к молекулам межклеточной адгезии лимфоцитов или эндотелия либо растворимыми препаратами самих этих молекул. Именно на таком подходе основан новый способ лечения болезней иммунологического патогенеза.
Нормальная миграция лейкоцитов. Oписанные выше молекулы, регулирующие миграцию клеток в область воспаления, необходимо отличать от молекул, ответственных за нормальную миграцию лимфоцитов. Непримированные лимфоциты экспрессируют L-селектин, обеспечивающий связывание с углеводными лигандами на эндотелии ВЭВ (и принимающий участие в прилипании к эндотелию) в лимфоидных образованиях слизистых оболочек и в периферических лимфоузлах. Останавливаясь, например, в пейеровой бляшке, они могут связываться при участии интегрина α4β7 с МАdСАМ-1 на поверхности эндотелия. Поскольку интегрин α4β7 обеспечивает миграцию лимфоцитов в лимфоидную ткань слизистых оболочек, а интегрин α4β1 — связывание с VCAM-1 на активированном эндотелии или с фибронектином во внеклеточном матриксе, экспрессия той или другой из этих молекул означает различное направление миграции — непримированных лимфоцитов в нормальную лимфоидную ткань или активированных Т-клеток в очаг воспаления.
Взаимодействие лейкоцитов с внеклеточным матриксом. Пройдя сквозь эндотелий сосуда и проникнув в ткань, лейкоциты неминуемо должны взаимодействовать с белками внеклеточного матрикса (коллаген, ламинин, фибронектин и др.), а также с клетками ткани. Покинувшие кровеносный сосуд лимфоциты сразу теряют в результате энзиматического расщепления уже ненужный им L-селектин. Происходит смена функционального фенотипа: циркулировавшая клетка становится приспособленной к перемещению в тканях.
Многие из молекул лейкоцитарной поверхности, обеспечивающих взаимодействие с внеклеточным матриксом, входят в группу β1-интегринов; они названы «очень поздними антигенами» (verylate antigens, VLA), поскольку впервые были идентифицированы на поверхности Т-клеток в поздней стадии активации. Теперь все β1-интегрины называют VLA, хотя большинство их присутствует не только на лимфоцитах. Эта группа включает рецепторы для коллагена (VLA-2 и VLA-3), ламинина (VLA-3 и VLA-6) и фибронектина (VLA-3, VLA-4 и VLA-5). Сам факт появления некоторых из этих молекул, означающий отдаленный результат активации лимфоцитов, свидетельствует о выполнении клетками некой программы дифференцировки, в которой взаимодействие с внеклеточным матриксом — эго один из наиболее поздних этапов.
Хемотаксические молекулы стимулируют лейкоциты к миграции и определяют ее направление
Интегрины, с помощью которых лейкоциты проникают сквозь эндотелий, присутствуют на клеточной поверхности или, прежде чем попасть на нее, сохраняются во внутриклеточных гранулах; при этом большинство из них неактивно и для функционирования нуждается в исходящем от эндотелия сигнале активации. Сигнальные молекулы могут быть продуктами самого эндотелия или осевшими на нем пептидами, которые выделяет подлежащая ткань. Многие из сигнальных молекул обладают также хемотаксическими свойствами; в том числе это С5а, лейкотрнен-В4 и разнообразные низкомолекулярные цитокины, получившие общее название «хемокины» (рис. 5.12).
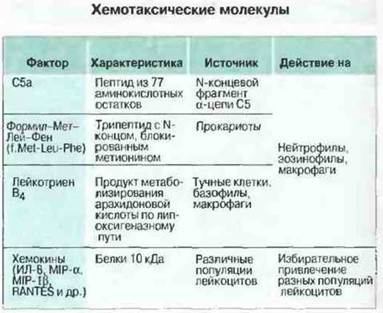
Рис. 5.12. Молекулы, вызывающие хемотаксис разных популяций лейкоцитов в зависимости от экспрессии на их поверхности специфических рецепторов. Некоторые из этих хемокинов оказывают на клетки хемотаксическое действие, другие - только активирующее.
Хемотаксическую активность клеток следует отличать от хемокинетической: хемотаксис - это направленная миграция клеток по градиенту концентрации хемотаксических молекул, а хемокинез — беспорядочное перемещение клеток. В основе направленной миграции лежит способность клетки отвечать на градиент концентрации хемотаксического медиатора при величине этого градиента между ее ведущим и концевым полюсами не менее 0,1%. Хемокинез связан с усилением общей подвижности клеток под действием того или иного медиатора, например гистамина.
Хемокины. Это группа хемотаксических гепарин-связывающих молекул, в которую входят не менее 25 низкомолекулярных цитокинов, в частности ИЛ-8 и RANTES1. Хемокины высвобождаются в очаге воспаления и могут связываться на поверхности эндотелия, взаимодействуя с сульфатными группами присутствующего на нем гепарина. Многие из этих цитокинов связывает также антиген DARC — групповой антиген крови системы Даффи, избирательно экспрессируемый на эндотелии венул. Связанные с поверхностью эндотелия хемокины могут вызывать повышение авидности интегринов на лейкоцитах в фазе «краевою стояния», остановленных при участии селектинов. Большинство хемокинов синтезируется лейкоцитами, однако ИЛ-8 и хемотаксический для макрофагов белок 1 (МСР-1. от англ. macrophage chemotactic protein-1) продуцирует, например, культура клеток эндотелия, причем активация этих клеток цитокинами, способствующими развитию воспаления, усиливает синтез. Хемокины и другие хемотаксические молекулы, рассматриваемые ниже, воздействуют на клетки посредством «змеевидных» рецепторов, обладающих семью трансмембранными сегментами. Различные змеевидные рецепторы (разной специфичности) избирательно распределены среди отдельных популяций лейкоцитов, чем отчасти можно объяснить избирательность действия различных хемокинов, например тот факт, что макрофагальный воспалительный белок Iβ (MIP- Iβ) стимулирует исключительно Т-клетки CD8+. Некоторые хемокины только активируют клетки, другие проявляют в первую очередь хемотаксические свойства, третьи сочетают обе функции. С чем связана необходимость!!таком разнообразии хемокинов, неясно, но можно предполагать, что за счет этого разнообразия возможна избирательная регуляция перемещения лейкоцитов как у поверхности эндотелия, так и в тканях.
Другие хемотаксические молекулы. Ряд белков вызывает хемотаксис нейтрофилов и макрофагов (рис. 5.12). Эти клетки имеют рецепторы для пептидов, блокированных на N-конце формилметионином, в частности рецептор, связывающий трипептид f.Met-Leu-Phe (f.MLP). Поскольку при трансляции всех белков у прокариот в отличие от эукариот инициаторной аминокислотой служит метионин, он и выполняет роль простого специфического сигнала присутствия бактерий, по направлению к которым должны устремиться фагоциты. Нейтрофилы и макрофаги имеют также рецепторы для С5а и лейкотриена В4. Оба эти хемоаттрактанта образуются в очаге воспаления: С5а — в результате активации комплемента, лейкотриен LTB4 — при активации разнообразных клеток, чаше всего макрофагов и тучных. Кроме того, хемотаксис фагоцитов вызывают молекулы, образуемые системой свертывания крови, прежде всего фибриновый пептид В и тромбин.
Клетки, прибывшие в очаг воспаления первыми, способны в результате активации вызвать следующую волну лейкоцитарной миграции. Так, активированные моноциты выделяют ИЛ-8, который может вызвать хемотаксис нейтрофилов и базофилов. Подобно этому, активация макрофагов приводит к метаболизированию арахндоновой кислоты с образованием и выделением лейкотриена В4.
1RANTES - Regulated on Activation. Normal T-cell Expressed and Secreted - хемокин, выделяемый неиммунными Т-клетками при активации; мол. масса 7,8—8,7 кДа; хемоаттрактант для моноцитов и Т-клеток CD4+/CD45RO+. — Прим. перев.