ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 9. Распознавание антигена
СПЕЦИФИЧНОСТЬ И АФФИННОСТЬ АНТИТЕЛ
Реакциям антиген-антитело свойственна высокая специфичность. Например, противокоревые антитела связываются с вирусами кори и создают иммунитет к этому заболеванию, но не способны связаться с вирусами других видов, в частности с вирусами полиомиелита, и не защищают от них организм. Специфичность антисыворотки суммарно отражает специфичность содержащихся в ней антител, в популяции которых может присутствовать множество паратопов, способных связываться с различными эпитопами или даже с разными частями одного и того же эпитопа (рис. 9.8). Однако, если антиген А имеет общие эпитопы с антигеном Б, часть антител, специфичных к А, будет реагировать также и с Б. Этот феномен назван перекрестной реактивностью.
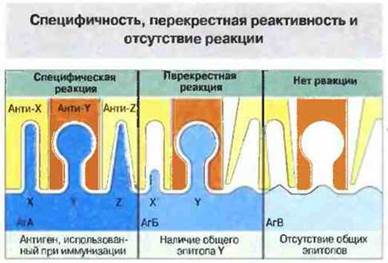
Рис. 9.8. Специфичность антисыворотки отражает специфичность содержащихся в ней отдельных молекул антител (например, антител анти-Х, анти-Y и анти-Z к эпитопам X, Y и Z, встречающимся на разных антигенных молекулах). Антиген А (АгА) и антиген Б (АгБ) обладают общим эпитопом Y. Антисыворотка против АгА (анти-XYZ) не только специфически связывается с АгА, но и перекрестно реагирует с АгБ (благодаря распознаванию эпитопов Y и X'), но не дает реакции с АгВ, поскольку он не имеет общих эпитопов с АгА.
Антитела распознают наружную конформацию антигенов
Антитела распознают, разумеется, не отдельные химические группы, а пространственную форму эпитопов (рис. 9.9), причем с поразительной специфичностью, улавливая, кроме различий в распределении зарядов, в оптической и стереоизомерии, также и минимальные различия в первичной аминокислотной последовательности (в случае белковых антигенов) (рис. 9.10). Вследствие столь тонкой специфичности большая часть антител способна связываться только с нативными (не денатурированными) антигенами или с такими фрагментами антигена, которые сохраняют третичную структуру, необходимую для множественных взаимодействий при образовании связи между паратопом и эпитопом.
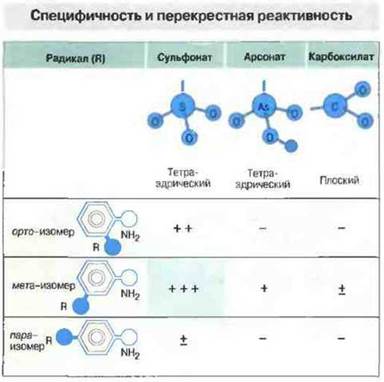
Рис. 9.9. Антисыворотка, полученная при иммунизации животных мега-аминобензолсульфонатом в качестве гаптена, была исследована в реакции связывания с орто- и пара-изомерами того же гаптена, а также с тремя изомерами (орто-, мета- и пара-) двух различных, но химически родственных антигенов - аминобензоларсоната и аминобензолкарбоксилата. Антисыворотка специфически реагирует с гаптеном, сульфонатная группа которого находится в мета-положении, но дает также перекрестную (хотя и слабую) реакцию с ортоизомером того же гаптена. Еще более слабые перекрестные реакции наблюдаются с гаптеном, имеющим арсонатную или карбоксилатную группу в мета-, но не в орто- или пара-положении. В отличие от сульфонатной группы, арсонатная группа крупнее по размерам и имеет дополнительный атом водорода, тогда как карбоксилатная группа меньше той и другой. Эти данные свидетельствуют о том, что для связывания антител с антигеном важна не только его конфигурация, но и отдельные содержащиеся в его молекуле химические группы.
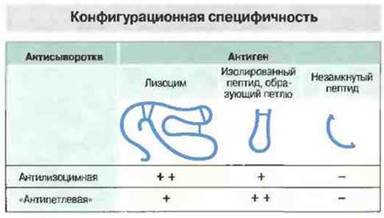
Рис. 9.10. В молекуле лизоцима имеются внутрицепочечные дисульфидные связи (показаны красными полосками), благодаря которым полипептид образует петли. Антисыворотки к целой молекуле лизоцима (антилизоцимная) и к одной из пептидных петель («антипетлевая») различают оба эти антигена, но не реагируют с тем же пептидом в незамкнутой (восстановленной) форме. Этим подтверждается значение третичной структуры антигена как фактора, определяющего специфичность антител.
В связи с этим в иммунологических исследованиях при получении специфических антител могут возникать затруднения. Для упрощения работы часто в качестве антигена используют специально синтезированный короткий полипептид известной первичной структуры, поскольку это легче, чем путем очистки получить достаточное количество нативного антигена. Однако антитела, образующиеся в результате иммунизации синтетическими пептидами, часто не обладают требуемой специфичностью и аффинностью к антигену в его нативной форме.
Модифицируя гены, можно получать антитела заданной конструкции
Благодаря достижениям молекулярной биологии стало возможным получать моноклональные антитела (мАт) заданной специфичности, аффинности и изотипа и, более того, конструировать гены антител, содержащие V-гены мыши и С-гены человека. Иммунизируя мышей, можно вызвать образование антител нужной специфичности, а затем придать им видовую антигенностъ антител человека. Благодаря весьма низкой чужеродности таких мАт для человека они могут использоваться в клинике. Еще более изощренные методы конструирования позволяют дополнять каркас V-домена антител, получаемых от животных одного вида, набором участков, определяющих комплементарность (CDR, см. гл. 8), из антител животных другого вида. Можно даже производить замены определенных аминокислотных остатков в антигенсвязывающем центре для увеличения аффинности антител (рис. 9.11).
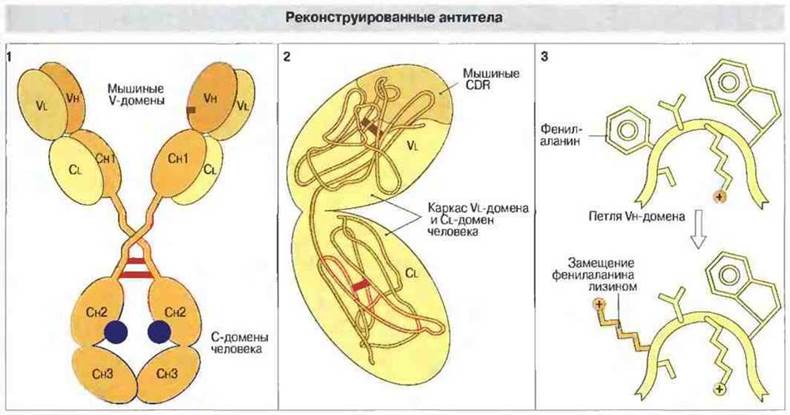
Рис. 9.11. Мышиные V-гены можно соединить с С-генами человека и ввести продукт рекомбинации в геном клетки, которая в результате будет синтезировать химерные «мышино-человеческие» антитела (1) Другой вариант конструирования антител заключается в присоединении к каркасу V-области легкой цепи гипервариабельных участков (CDR) нужной структуры (2). Кроме того, аффинность антигенсвязывающего центра можно изменить, вызвав точечную мутацию в том участке ДНК, который кодирует гипервариабельные петли Vн-домена (в данном случае показана замена фенилаланина на лизин) (3).
Полифункциональные антигенсвязывающие центры могут быть одновременно комплементарны различным эпитопам
Не так давно было обнаружено, что одно и то же антитело может быть комплементарно к нескольким различным антигенным детерминантам. Они конкурируют за связывание с этим антителом и, по-видимому, занимают различные участки полости, образуемой антигенсвязывающим центром (рис. 9.12).
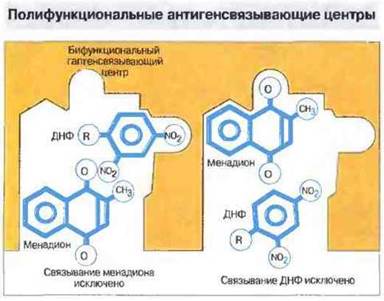
Рис. 9.12. Один и тот же антигенсвязывающий центр способен соединяться с несколькими разными антигенными детерминантами. Например, в антигенсвязывающем центре антител 460 имеются две ниши, расположенные на расстоянии 1,2-1,4 нм, одна из которых связывает менадионовую (витамин К3), а другая - динитрофенильную (ДНФ) гаптенную группу. Это связывание происходит конкурентно, т. е. связывание одного гаптена исключает последующее связывание другого. Антигенсвязывающие центры, способные взаимодействовать с несколькими антигенными детерминантами, названы полифункциональными.
Специфичность популяции поликлональных антител к данному антигену (А) не означает, что каждый вид входящих в нее антител взаимодействует исключительно с А. Просто большая часть этих антител обладает полифункциональным активным центром, способным связывать А, и суммарная реактивность в отношении А существенно превышает реактивность в отношении прочих антигенов. Иными словами, специфичность (поликлональных) антител может быть популяционным феноменом — усредненной характеристикой всех антител, содержащихся в антисыворотке (рис. 9.13).
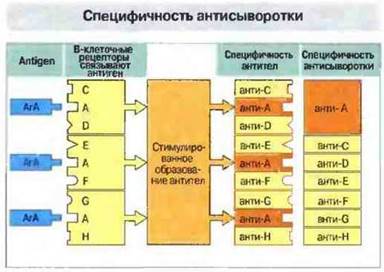
Рис. 9.13. Специфичность антисыворотки как популяционный феномен. Отдельно взятый антиген А (АгА) может связаться с рецепторами разных В-клеток, специфичными не только к А, но и к другим антигенам (С, D, Е, F ит. д.), т. е. обладающими полифункциональными активными центрами. Каждая В-клетка, активированная АгА, образует антитела, специфичные не только к АгА, но и к другим антигенам Однако благодаря тому, что все активированные В-клетки специфичны к АгА, но не все из них специфичны к другим антигенам, концентрация антител к АгА будет высокой, а к другим антигенам - низкой.
Высокоаффинные антитела часто эффективнее низкоаффинных
Аффинность связывания антител с антигеном представляет не только теоретический интерес. От нее, как и отавидности, зависят физиологические и патологические эффекты антител. Высокоаффинные антитела гораздо активнее низкоаффинных в ряде биологических реакций (рис. 9.14). В экспериментах на животных иммунные комплексы, содержащие низкоаффинные антитела, длительно персистируют в циркуляции и откладываются на базальной мембране капилляров почек, нарушая их функцию. В то же время иммунные комплексы, образованные высокоаффинными антителами, намного быстрее выводятся из кровотока и локализуются в мезангии почечных клубочков, мало влияя на их функцию.

Рис. 9.14. Биологические реакции, в которых высокоаффинные антитела эффективнее низкоаффинных.
Аффинность антител к большинству Т-зависимых антигенов нарастает в ходе иммунного ответа. Ее увеличения можно достичь также особыми способами иммунизации. Например, субпопуляция высокоаффинных антител в сыворотке увеличится, если антиген вводить вместе с ИФγ (рис. 9.15). Ряд иммуноадъювантов стимулирует образование антител, но лишь некоторые способны повысить их аффинность. Поскольку от этого свойства зависит эффективность антител, ИФγ вполне пригоден для использования в качестве адъюванта в составе вакцин.
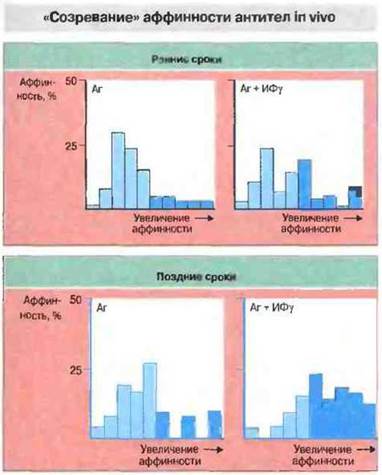
Рис. 9.15. Мышей опытной группы иммунизировали антигеном вместе с 30 000 ЕД ИФγ (Аг + ИФγ), а контрольной - только антигеном (Аг). Аффинность образующихся антител определяли на ранних и поздних сроках после иммунизации. В сыворотке мышей опытной группы независимо от сроков взятия крови выявлялось больше высокоаффинных антител (темно-голубые столбики), чем в сыворотке контрольной группы. (По Holland, Holland and Steward, 1990. Clin. Exp. Immunol. 82: 221-226.)
Определение аффинности антител
Существует ряд методов определения аффинности антител и авидности антисывороток. Процедура каждого из них заключается в том, чтобы создать условия, в которых взаимодействие антител с антигеном достигает равновесия: Ат + Аг ⇔ АтАг, а затем, не нарушая этого равновесия, измерить концентрации свободного и связанного антигена. Сделать это можно по-разному, например отделяя свободный антиген от связанного с помощью таких методов, как диализ, гель-фильтрация, центрифугирование или избирательное осаждение, либо определяя изменения флуоресценции антигена или антител после образования иммунного комплекса.
Используя полученные данные для расчета по формуле закона действующих масс, можно вычислить константу равновесия (аффинности), К:
![]()
где [АтАг] — концентрация иммунных комплексов, (Аг) — концентрация свободного антигена и [Ат] — концентрация незанятых антигенсвязывающих центров в условиях равновесия. Если половина антигенсвязывающих центров связана с антигеном и, следовательно, [Ат] равна [АтАг], значение К равно 1/[Аг]. Иными словами, для связывания 50 % активных центров в случае высокоаффинных антител (т. е. имеющих высокую К) достаточно низкой концентрации антигена, а в случае низкоаффинных антител нужна его высокая концентрация.
Распределение содержащихся в антисыворотке антител по аффинности отличается от нормального
В реакциях взаимодействия фермента с субстратом константа равновесия (К) неизменна при различных концентрациях субстрата. При связывании же антител с гомологичным антигеном дело обстоит иначе - популяция поликлональных антител гетерогенна по этой константе, т. е. по величине аффинности. Ранее предполагалось, что в любой антисыворотке количественное распределение антител по аффинности имеет характер нормального (Гауссова) распределения. Как теперь установлено, это предположение было ошибочным. Графический анализ данных по аффинности для реакции между антисывороткой и антигеном показывает, что распределение антител по аффинности не соответствует нормальному (рис. 9.16). Однако среднюю аффинность популяции антител в данной антисыворотке (К0) можно определить с хорошим приближением как величину, обратную такой концентрации свободного антигена, которая необходима для насыщения половины всех антигенсвязывающих центров антител: К0 = 1/[Агсвоб.].
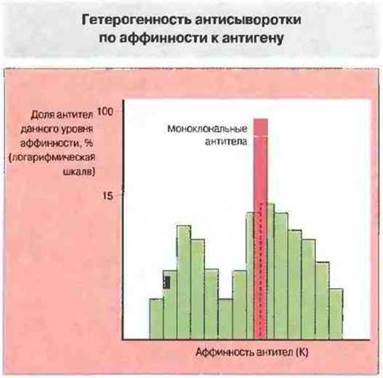
Рис. 9.16. Гистограмма типичного распределения антител по аффинности в антисыворотке к данной антигенной детерминанте в сравнении с однородной по аффинности популяцией моноклональных антител к той же детерминанте. В отличие от поликлональных все моноклональные антитела характеризуются одинаковой аффинностью. Распределение поликлональных антител, содержащихся в антисыворотке, по аффинности отличается от нормального.
Гетерогенность популяции антител по аффинности трудно оценить экспериментальными методами. Гетерогенность популяции антител по аффинности невозможно точно описать математической формулой, поскольку нельзя с уверенностью предполагать характер их распределения по этой величине в каждой отдельной антисыворотке — соответствует ли он нормальному, асимметричному, би- или полимодальному. Для определения такой гетерогенности используется довольно сложный компьютерный анализ кривых связывания, основанный на допущении, что распределение соответствует нормальному. Описан, однако, и простой экспериментальный подход для ее оценки, не основанный па таком допущении. Согласно этой методике, в панели для микротитрования, лунки которых предварительно нагружены. т. е. покрыты, иммобилизованным антигеном, вносят раствор исследуемых антител в известной концентрации и серийные разведения (растворы последовательно уменьшающихся концентраций) свободного антигена, подавляющего связывание антител с антигеном, иммобилизованным на поверхности лунок. Затем рассчитывают в процентах подавление связывания для каждого разведения свободною антигена и определяют молярную концентрацию, обеспечивающую 50 % подавление (I0.5). Выразив результаты в процентилях, строят гистограмму и анализируют по ней разброс значений аффинности. Математической величиной для его выражения может служить индекс Шаннона, высокие значения которого свидетельствуют о широком диапазоне аффинности в популяции антител, а низкие соответственно об узком диапазоне.