ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 11. Взаимодействие клеток при гуморальном иммунном ответе
ВНУТРИКЛЕТОЧНЫЕ СИГНАЛЫ ПРИ АКТИВАЦИИ ЛИМФОЦИТОВ
Комплекс ТкР способен передавать сигналы внутрь клетки. В передаче сигналов участвуют комплекс CD3 (γ-, δ-, ε-, ζ- и η-цепи) и фермент р56lск, который связан с внутриклеточными участками молекул CD4 или CD8 (р56lск - сокращенное название специфичной для лимфоцитов тирозинкиназы с мол. массой 56 кДа). В-клетки также экспрессируют семейство молекул, участвующих в передаче сигналов: оно включает Igα, Igβ и мембраносвязанные IgM и IgD (см. гл. 6).
Активацию лимфоцитов инициируют тирозинкиназы
Распознавание Т-клеточным рецептором комплекса антиген—МНС-молекула и связывание костимулируюших молекул вызывают передачу сигнала внутрь Т-клеток. Самый первый внутриклеточный этап сигнализации состоит в фосфорилировании тирозина, осуществляемом с участием тирозинкиназ семейства srс, в частности lck, ассоциированной с CD4, и fyn; обе они фосфорилируют последовательности-мишени, обнаруженные в ζ-цепи CD3 и присутствующие также в молекулах Igα, Igβ и FcγR. Эти последовательности получили название иммунорецепторные тирозин-активируемые мотивы, ITAM (от англ. immunorecеptor tyrosine-based activation motifs). Тирозинкиназа ZAP-70 связывается с ITAM и активируется, в свою очередь активируя фосфолипазу С и тем самым открывая классический путь сигнализации. Сигналы от ТкР и CD28 интегрируются (рис. 11.12), активируя находящиеся в латентном состоянии цитоплазматические факторы транскрипции, такие как NF-AT или NF-kB, которые после этого мигрируют в ядро. Здесь они действуют на гены, необходимые для Т-клеточной активации, включая гены ИЛ-2 и рецептора ИЛ-2. Продуцируемый в результате этой активации ИЛ-2, связываясь со своим рецептором, вызывает деление клеток.
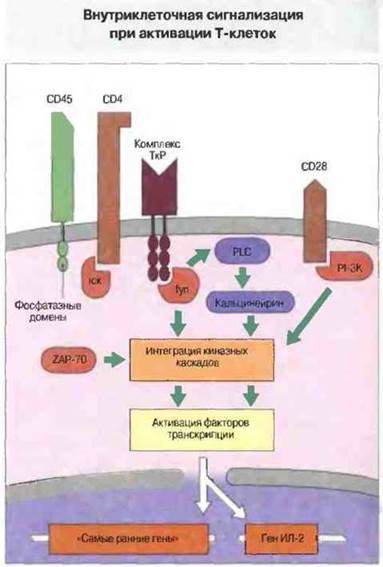
Рис. 11.12. При активации Т-клеток происходит передача сигналов от Т-клеточного рецептора и от молекулы CD28. Корецептор CD4, связанный с комплексом ТкР, взаимодействует с киназой lсk. Подобные киназы активируются в результате дефосфорилирования, возможно осуществляемого фосфатазными доменами общего лейкоцитарного антигена CD45. Активированная киназа lсk фосфорилирует домены ITАМ ζ-цепей CD3, что позволяет им ассоциировать с другими киназами, включая fyn и ZAP-70. Киназа fyn активирует фосфолипазу С (PLC), которая косвенным образом вызывает высвобождение внутриклеточного кальция. Кальций связывается с кальцинейрином и активирует факторы транскрипции. Одновременно с этим киназы ZAP-70, fyn и PI-3 (РI-3К, ассоциированная с CD28) передают интегрированные сигналы через киназные каскады в цитоплазме; эти сигналы также активируют факторы транскрипции. Поступая в ядро, эти факторы активируют гены, в том числе «самые ранние» гены клеточного деления, и ген ИЛ-2.
Процессы активации В- и Т-клеток протекают сходным образом
Функцию Т-клеточной молекулы CD3 у В-клеток выполняют молекулы Igα и Igβ, которые также содержат ITAM в своей цитоплазматической части. Перекрестное связывание поверхностных Igведет к активации киназ семейства srс, которые в В-клетках представлены не только fyn, но также lyn и blk. Киназа Syk, аналогичная киназе ZAP-70 Т-клеток, связывается с фосфорилированными Igα и Igβ. Это приводит к активации каскада киназ и переносу ядерных факторов транскрипции (аналогично происходящему в Т-клетках) (рис. 11.13).
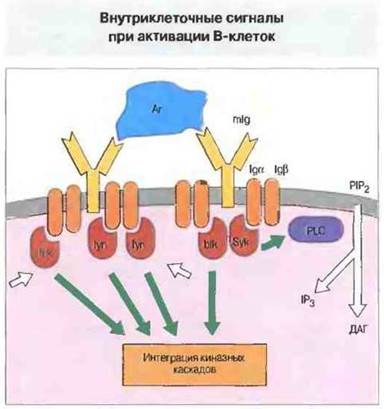
Рис. 11.13. Активация В- и Т-клеток происходит сходным образом. При перекрестном связывании мембраносвязанных Ig (mlg), например Т-независимым антигеном (Аг), активируются тирозинкиназы, в том числе lсk, lyn, fyn и blk. Они фосфорилируют домены ITAM в Igα- и Igβ-цепях рецепторного комплекса, которые приобретают способность связывать другую киназу, Syk, в свою очередь активирующую фосфолипазу С. Последняя действует на мембранный PIP2, в результате чего образуются IP3 и диацилглицерол (ДАТ), который активирует протеинкиназу С. Сигналы, передаваемые другими киназами, активируют ядерные факторы транскрипции, (mIg - мембранный Ig.)
В-клеточную активацию заметно усиливает «корецепторный комплекс», состоящий из 3 белков — рецептора 2 комплемента (CR2, CD21), CD19 и молекулы-мишени для антипролиферативных антител (ТАРА-1; CD81) (рис. 11.14). Известно, что фолликулярные дендритные клетки длительное время удерживают на своей поверхности антиген в виде иммунных комплексов (иккосом). Антиген в составе такого комплекса может связываться и с поверхностным Ig В-клеток, и (посредством соединенного с ним компонента C3d комплемента) с CR2. Это связывание ведет к фосфорилированию цитоплазматической части молекулы CD19, которая связывается также с киназами. Возможно, киназы, находясь близко к В-клеточному рецептору, способствуют активации В-клетки, особенно в тех случаях, когда концентрация антигена невелика.
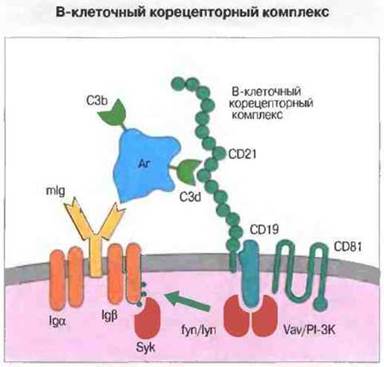
Рис. 11.14. В-клеточный корецепторный комплекс состоит из CD21 (рецептор комплемента 2 типа), CD19 и CD81 - молекулы с 4 трансмембранными сегментами. Антиген (Аг) с ковалентно присоединенным С3bили C3d может перекрестно связывать мембранный Ig (mIg) с CD21 корецепторного комплекса Это значительно снижает потребность клетки в антигене для ее акгивации. Молекула CD19 может взаимодействовать с киназами, включая lyn, fyn, Vav и киназу PI-3 (PI-3K. ср. с CD28 Т-клеток). Перекрестное связывание рецепторов вызывает фосфорилирование Igα- и Igβ-цепей комплекса антиген-рецептор, а также мобилизацию и активацию Syk.
Сигнал активации - это суммарный сигнал от антигенного рецептора и костимулирующих молекул
До сих пор не известно точно, что представляет собой эффективный антигенный сигнал. В случае Т-клетки взаимодействия антигена с одиночным ТкР недостаточно для активации; необходимое для стимуляции число таких взаимодействий может зависеть от наличия других стимулирующих сигналов, типа Т-клеток и степени их активации. Установлено, что легко активируются клетки мышиных Т-клеточных гибридом (полученных путем слияния нормальных Т-лимфоцитов и клеток Т-клеточной опухоли). Чтобы стимулировать гибридную клетку, эффективно функционирующая АПК (например, макрофаг) должна нести не менее 60 комплексов, образованных молекулой МНС класса II и антигенным фрагментом. Функционально менее активные АПК, такие как фибробласты с трансфицированным МНС-геном класса II, должны иметь для этого 5000 таких комплексов.
Проведенные недавно исследования показали, что активация требует взаимодействия примерно 8000 молекул ТкР с комплексами МНС—пептид; именно такое число ТкР исчезает с поверхности клетки при активации Т-клеточных клонов. Поскольку взаимодействие ТкР-МНС характеризуется низкой аффинностью, единичный комплекс МНС—пептид способен, возможно, активировать большое число ТкР. Не исключено, что для трансформированных Т-клеток, какими являются гибридомы, необходима менее сильная ТкР-активация. При наличии костимулирующего сигнала (например, связывания CD28/B7) для активации Т-клеточных клонов достаточно 1500 активированных и интернализованных ТкР.
Сведения подобного типа о действии Т-зависимых антигенов на В-клетки отсутствуют. Связывание Т-независимого антигена с одиночным рецептором не ведет к активации В-клетки, однако пока еще нет данных, которые позволили бы сопоставить эффекты стимуляции В-лимфоцитов с описанными выше процессами в Т-клетках.
Взаимодействие антигена с ТкР иди мембранным иммуноглобулином само по себе не может служить положительным сигналом активации для Т- или В-клеток. Возможно, его достаточно для генерации «отрицательного», или толерогенного, сигнала, однако и это сомнительно. В настоящее время считается, что для активации лимфоидных клеток необходим целый ряд взаимодействий, каждое из которых потенциально несет сигнальную функцию. Костимулирующие молекулы, например CD2 и CD11а/CD18, ответственны не только за связывание; их цитоплазматические домены участвуют в передаче сигналов. Так, эксперименты с удалением цитоплазматического домена CD2 показали, что в результате такого вмешательства процесс активации нарушается, однако адгезивная функция молекулы остается неизменной.
Митогены и суперантигены также могут активировать лимфоциты
Для изучения Т- и В-клеточной активации наряду с антигенами используются также митогены - вещества, которые в отличие от антигенов могут активировать Т- и В-клетки неспецифически. Для большинства Т-клеток стимуляторами служат фитогемагглютинин (ФГА), выделенный из красной фасоли, и конкапавалин А (КонА), экстрагированный из плодов клещевины. Механизм действия этих агентов детально изучен; они связываются с поверхностными молекулами Т-клеток, ответственными за активацию, в частности с ТкР и CD2.
Способностью неспецифически активировать Т-клетки обладают и молекулы другой группы, так называемые «суперантигены», большинство которых имеет бактериальное происхождение. К ним относятся стафилококковые энтеротоксины (вызывающие острые пищевые отравления некоторых типов), токсин, вызывающий развитие синдрома токсического шока при сепсисе, токсин эксфолиативного дерматита и некоторые вирусные белки. Суперантигены связываются с молекулами МНС класса II на АПК и распознаются ТкР, однако не посредством того же механизма, какой действует при распознавании Т-клеточным рецептором комплекса МНС—антигенный пептид. Супераптиген связывается только с Vβ-цепью ТкР, однако этого достаточно для активации Т-клетки (рис. 11.15). Эффект стимуляции при этом такой же, как и при использовании антигенов: в зависимости от условий эксперимента может возникнуть либо иммунный ответ, либо клональная анергия.
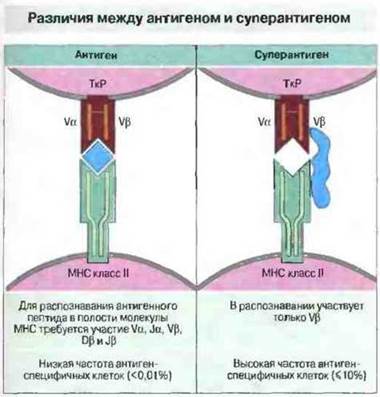
Рис. 11.15. Для активации ТкР антигенные пептиды, как правило, должны быть процессированы. Однако суперантигены, такие как стафилококковые энтеротоксины, не подвергаются процессингу, а непосредственно связываются с молекулой МНС класса II и Vβ-цепью ТкР. Каждый суперантиген активирует отдельную популяцию Т-клеток, экспрессирующих Vβ, в зависимости от того, каким сегментом Vβ-гена Т-клетки кодируется ее рецептор.