ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 14. Иммунологическая толерантность
ЦЕНТРАЛЬНАЯ ТИМИЧЕСКАЯ ТОЛЕРАНТНОСТЬ К АУТОАНТИГЕНАМ
В тимусе происходит формирование Т-клеток из клеток-предшественников с еще неперестроенными генами Т-клеточных рецепторов (ТкР). В процессе развития лимфоцитов в тимусе эти гены подвергаются перестройке, после чего Т- клетки начинают экспрессировать ТкР, способные распознавать продукты деградации антигенов или пептиды в связывающей их полости молекул МНС - антигенов, кодируемых генами главного комплекса гистосовместимости (МНС, от англ. major histocompatibility complex) организма (см. гл. 7 и 9).
В тимусе происходит отбор Т-клеток с рецепторами, способными связывать антигены в комплексе с молекулами МНС, и делеция клеток, высокоавидных к собственным антигенам
Высокая скорость пролиферации тимоцитов сочетается с их массовой гибелью: подавляющее большинство так называемых дважды положительных (CD4+CD8+) тимоцитов погибает в тимусе. К причинам этого относятся аберрантная перестройка генов ТкР (возникновение «неиспользуемых» рецепторов), отрицательная селекция и неспособность клеток пройти положительный отбор. Условием положительной селекции является умеренная степень авидности Т-клеток к полиморфным областям молекул МНС, ассоциированных с пептидами; такие клетки выживают (рис. 14.2). Они связываются с молекулами МНС на поверхности эпителиальных клеток корковой зоны, и это связывание предположительно защищает Т-клетки от запрограммированной гибели (апоптоза). Положительная селекция обеспечивает формирование зрелых Т-клеток, способных распознавать лишь те пептиды, которые удерживаются в специальной полости собственных молекул МНС. Этот феномен ограниченности распознавания называют рестрикцией по МНС. Однако положительная селекция не предотвращает дифференцировку Т-клеток, несущих рецепторы с высокой аффинностью и к собственным пептидам, и к молекулам МНС. Поэтому, чтобы заставить «молчать» такие высокоаутореактивные клетки, должен существовать тот или иной путь отрицательной селекции.
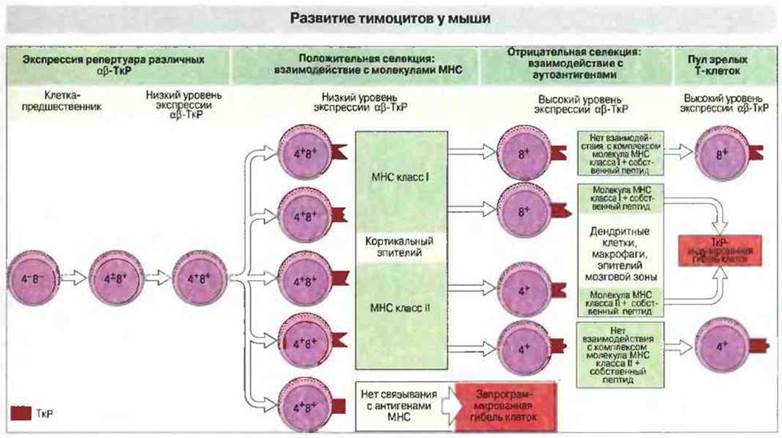
Рис. 14.2. Предшественники тимоцитов в корковой зоне тимуса превращаются в «дважды положительные» клетки, экспрессирующие незначительное количество αβ-ТкР. Эти клетки проходят положительную селекцию, взаимодействуя со «своими» молекулами МНС класса I или II на кортикальном эпителии. Неотобранные клетки (их большинство) погибают в результате запрограммированного саморазрушения (апоптоза). Положительно отобранные клетки теряют одну из своих корецепторных молекул (CD4 или CD8). И наконец, аутореактивные клетки элиминируются за счет их взаимодействия с собственными пептидами, презентированными на клетках в области кортико-медуллярного соединения и мозговой зоне тимуса.
Механизмы положительной и отрицательной селекции
Поскольку и положительная, и отрицательная селекция связаны с распознаванием собственных пептидов в ассоциации с собственными молекулами МНС, возникает вопрос, каким образом сигналы, передаваемые одним и тем же ТкР, обусловливают либо тот, либо другой процесс. Для объяснения этого предложены две теории.
✵ Согласно так называемой количественной теории, в случае незначительного числа комплексов пептид-молекула МНС индуцируется положительная селекция, тогда как при высоком их числе — отрицательная.
✵ Основное положение «качественной» теории состоит в том, что, воздействуя на один и тот же Т-клеточный рецептор, различные комплексы пептид-молекула МНС индуцируют сигналы разного типа, вызывая либо положительную, либо отрицательную селекцию.
В пользу количественной теории говорят результаты опытов на трансгенных мышах, экспрессирующих ТкР к основному белку миелина — аутоантигену миелиновой оболочки, окружающей аксоны. У мышей одной из линий естественный пептидный эпитоп, распознаваемый специфичными к этому белку Т-клетками, связывается соответствующей молекулой МНС класса II, но с очень низкой аффинностью. Вследствие этого специфичные к данному эпитопу Т-клетки избегают отрицательной селекции в тимусе. Однако, если трансгенным мышам, имеющим тот же самый ТкР, ввести высокоаффинный аналог этого пептида, происходит отрицательная селекция незрелых тимоцитов — апоптотическая гибель (рис. 14.3). Очевидно, судьбу клеток — гибель или выживание — определяет число комплексов пептид молекула МНС, презентированиых развивающимся Т-клеткам. Собственные эпитопы, формирующие более стабильные комплексы с белками МНС, с большей вероятностью индуцируют отрицательную селекцию.
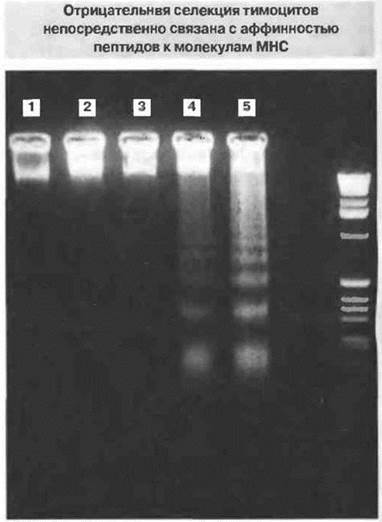
Рис. 14.3. Этот агарозный гель, окрашенный на ДНК, демонстрирует различную степень апоптоза (фрагментвции ДНК) тимоцитов трансгенных мышей (экспрессирующих ТкР, специфичные к пептидам Ас1-9 основного белка миелина) в присутствии пептидов с возрастающей аффинностью к молекулам МНС класса II.
1. Посторонний пептид.
2. Аутентичный низкоаффинный пептид (Ас1-9).
3. Аналоге промежуточной аффинностью к молекуле МНС (Ас1-9 [4А])
4. Аналог с высокой аффинностью к молекуле МНС (Ас1-9 [4V])
5. Другой аналог с высокой аффинностью к молекуле МНС (Ас1-9 [4Y]).
Качественная теория была предложена для объяснения результатов, полученных в опытах с органными культурами тимуса. Как показали эти эксперименты, пептидные эпитопы, измененные по аминокислотным остаткам, которые обеспечивают взаимодействие с Т-клеточным рецептором, — ТкР-антагонисты — вызывают более выраженную положительную селекцию, чем исходный эпитоп (агонист). Однако эти две теории не являются взаимоисключающими. Разработанные к настоящему времени методы позволяют измерять аффинность связывания очищенных молекул ТкР с очищенными молекулами МНС, нагруженными пептидными агонистами или антагонистами. При помощи этих методов установлено, что с теми комплексами пептид-антагонист-молекула МНС, которые индуцируют положительную селекцию в органных культурах тимуса, ТкР образует нестабильное соединение (рис. 14.4). Тот же самый ТкР относительно стабильно связывается с комплексом агонист—молекула МНС, и в тех же экспериментальных условиях это ведет к отрицательной селекции. Полученные результаты сближают количественную и качественную теории и соответствуют кинетической модели, согласно которой индукция процессов положительной или отрицательной селекции определяется скоростью диссоциации ТкР от комплекса пептид—молекула МНС. На общую авидность взаимодействия Т-клеток с антигенпрезентирующими клетками (АПК) в тимусе (рис. 14.5) влияют следующие факторы:
✵ число комплексов пептид-МНС; оно зависит от аффинности эпитопа к соответствующей молекуле МНС;
✵ число молекул ТкР и соответствующих корецепторных молекул, экспрессируемых Т-клеткой, и
✵ аффинность специфичного ТкР к лиганду.
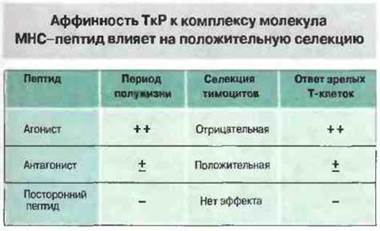
Рис. 14.4. Аффинность растворимого ТкР к комплексам из различных пептидов и соответствующих элементов рестрикции (антигены МНС) можно определить биофизическими методами, например методом поверхностного плазмонного резонанса. Существует прямая корреляция между временем полужизни ТкР-связи с комплексом молекула МНС-пептид и реакцией зрелых Т-клеток, экспрессирующих тот же рецептор (т. е. агонист > антагонист > посторонний пептид). Однако в органной культуре тимуса добавление пептида-агониста вызывает делецию развивающихся клеток (отрицательная селекция), тогда как пептид-антагонист стимулирует положительную селекцию. Это показывает, что низкоавидное взаимодействие стимулирует положительную селекцию, а высокоавидное - отрицательную.
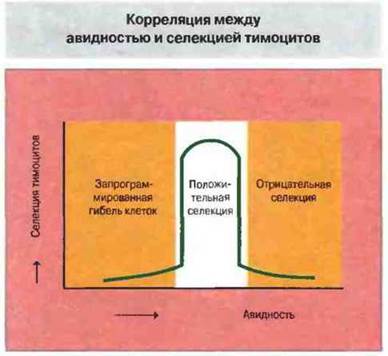
Рис. 14.5. Авидность взаимодействия Т-клетки с антигенным пептидом, презентированным на АПК, зависит от уровня экспрессии (концентрации) комплекса МНС-молекула-пептид [МНС-молекула + пептид] на АПК, а также от аффинности и экспрессии (концентрации) на поверхности Т-клеток ТкР и корецепторных молекул [ТкР + корецепторные молекулы]. [МНС-молекула + пептид] зависит от аффинности пептида к молекуле МНС и стабильности сформировавшегося комплекса. [ТкР + корецепторные молекулы] определяется аффинностью индивидуального ТкР к комплексу пептид- МНС-молекула, уровнем экспрессии ТкР на клеточной поверхности и соответствующим уровнем экспрессии корецепторных молекул.
Положительная селекция и коммитирование Т-клеток происходят на определенной стадии их развития
В связи с чем тимоциты в процессе своего развития утрачивают одну из корецепторных молекул и становятся либо CD4+-, либо С08+-клетками? Положительной селекции подвергаются лишь те незрелые тимоциты, которые имеют ТкР соответствующей специфичности и пока еще экспрессируют оба корецептора. Положительно отобранные тимоциты дифференцируются в так называемые моноположительные клетки, CD4+ или CD8+. При коммитировании в тот или иной тип клеток избирательно прекращается синтез яругой корецепторной молекулы. У мыши такое избирательное прекращение синтеза происходит, когда клетки, несущие оба корецептора, получают сигналы, необходимые для положительной селекции (рис. 14.2).
Время и место отрицательной селекции зависят от разнообразных факторов
В число таких факторов входят: доступность собственных антигенов для развивающихся Т-клеток, сочетанная авидность ТкР и вспомогательных молекул (CD8 или CD4) к комплексу собственный антиген МНС—собственный пептид и природа клеток, ответственных за делению. Отрицательная селекция не требует участия специализированных АПК; в обычных условиях это функция дендритных клеток тимуса или макрофагов, которые локализованы преимущественно в области перехода корковой зоны в мозговую; они несут на своей поверхности большое число молекул МНС классов I и II, обеспечивая тем самым связывание Т-клеток, имеющих высокую авидность к собственным пептидам (рис. 14.6). В отрицательной селекции могут также участвовать некоторые эпителиальные клетки мозговой и корковой зон. Делению способны вызывать и сами тимоциты (рис. 14.6).
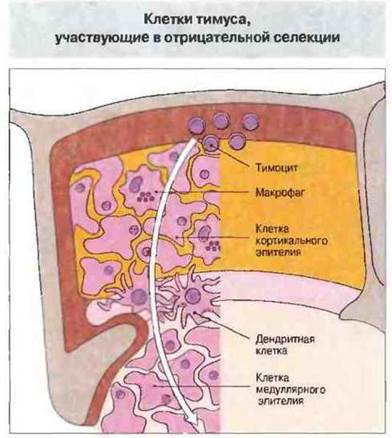
Рис. 14.6. Делегирующая популяция включает макрофаги костномозгового происхождения или дендритные клетки, расположенные преимущественно в области перехода корковой зоны в мозговую. В делеции могут участвовать и другие клетки, в частности сами тимоциты, обладающие «вето»-функцией, а также тимусные эпителиальные клетки некоторых типов, локализованные, возможно, в мозговой зоне.
В отрицательной селекции участвует специальный сигнал гибели
Существует ряд механизмов апоптотической гибели зрелых лимфоцитов. Сигналы, индуцирующие апоптоз, могут передаваться через Fas, CTLA-4 или рецепторы ФИО (см. ниже). Процесс отрицательной селекции дважды положительных тимоцитов (CD4+CD8+) уникален тем, что не зависит ни от одного из этих процессов.