Эволюция вирусов - Жданов В. М. 1990
Специальная часть
Коронавирусы
Коронавирусы образуют небольшую группу вирусов, поражающих млекопитающих и птиц, которые выделены в отдельное семейство Coronaviridae с одним родом Coronavirus [Закстельская Л. Я., 1982; Matthews R., 1982; Tyrrell D. et al., 1986].
Геном коронавирусов представляет собой однонитевую РНК с молекулярной массой 5,5х106—6,1х106, позитивной полярностью. На 3'-конце молекулы находится поли(А) - последовательность. На 5'-конце имеется кэп-структура. Вирионы имеют сферическую форму, диаметр 75—160 нм, характерные булавообразные выступы (12x24 нм) на внешней оболочке, образующие корону (откуда и название вирусов). Внутри внешней оболочки заключен нуклеокапсид, состоящий из рибонуклеопротеидов со спиральным типом симметрии, диаметром 11—13 нм (рис. 18). Вирионы имеют плотность 1,18 г/мл, размер 75—180 нм, состоят из сердцевины и внешней оболочки, содержат 4—6 белков, в их числе белки нуклеопротеида, матрикса и оболочечные гликопротеиды. Их молекулярная масса колеблется в пределах 15 000—200 000. Белок рибонуклеопротеида с молекулярной массой 50 000 образует тримеры с массой 140 000. Гемагглютинирующие свойства связаны с оболочечными антигенами; между вирусами внутри рода существуют серологические связи. В составе вирионов имеются фосфопротеид нуклеокапсида (50 000—60 000), гликопротеиды наружной оболочки E1 (20 000—35000) и Е2 (80 000—200 000), последний образует булавовидные выступы и обладает гемагглютинирующей активностью. На вирионной РНК гены расположены в следующем порядке: 5'- 200 000 NS — 35 000 NS — E2 — 14 000 NS — E1 —N-3'.
неструктурных белков 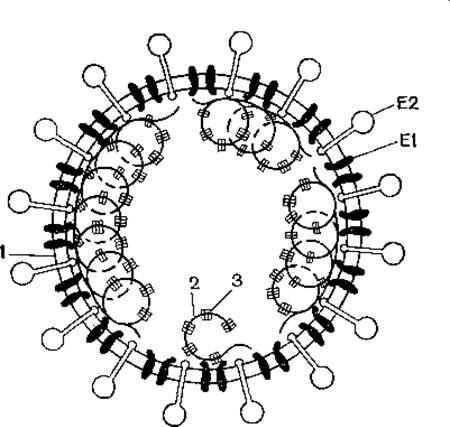
Рис. 18. Строение коронавируса (схема).
1 — липидный бислой; E1, Е2 — вирусные гликопротеиды, Е2 образует пепломеры, E1 взаимодействует с нуклеокапсидом; 2 - РНК; 3 — белки нуклеокапсида.
После прикрепления к рецепторам клеточной мембраны, эндоцитоза, слияния вирусной и клеточной мембран и депротеинизации геном коронавирусов начинает функционировать, индуцируя синтез белков (первичная транскрипция). При этом транслируется только часть генома, примыкающая к 5'-концу РНК и кодирующая синтез (200000). Этот пептид обладает полимеразной активностью. Вслед за этим синтезируется минус-нить, образуется репликативный интермедиат, на котором синтезируются дочерние нити РНК. Минус-нити являются матрицей для синтеза не только дочерних геномных РНК, но и 6 субгеномных РНК с молекулярной массой от 0,6х106 до 3,7х106. Все эти мРНК синтезируются с одной и той же точки отсчета на 3'- конце минус-нити, причем каждая из длинных нитей включает предыдущую, более короткую. Однако трансляция их происходит только из прилегающей к 5'-концу части. У 5'-конца геномной РНК имеются кэп-структуры, прикрепленные к лидерной последовательности длиной около 70 нуклеотидов. Эта последовательность находится у всех 6 мРНК, формируясь, по-видимому, посредством сплайсинга. Субгеномные РНК имеют кэп-структуры и поли (А)-последовательность. На рис. 19 приведена схема синтеза этих РНК коронавирусов. Участки, начинающиеся от 5'-концов, гомологичны. Особенности репликации коронавирусов — синтез субгеномных РНК разных размеров, начинающихся с одной и той же точки, — являются причиной высокой частоты получения рекомбинантов при заражении клеток двумя разными штаммами вируса мышиного гепатита [Makino S. et al., 1986].
В клетке «хозяина» синтезируются 6 вирионных белков: большой гликопротеид (gp84/90), малый гликопротеид gp31, фосфопротеид нуклеокапсида р51, два других мажорных белка р36 и р23 и минорный белок р28. При иммунологическом анализе белков коронавирусов, выделенных от разных животных, были получены следующие данные. Белки коронавирусов коров gp120, gp120, р52 и gp26 оказались иммунологически родственными сходным белкам коронавируса человека, но только трем белкам коронавируса мышей (вирусы гепатита): gpl90, р52, gp26 [Hogne В. et al., 1984]. Коронавирусы вызывают у человека острые респираторные заболевания и энтериты. Коронавирусы у свиней вызывают гастроэнтериты и энцефалит, у телят, собак и кошек — кишечные заболевания, у крыс — поражения дыхательных путей, у мышей — гепатит. Птичьи коронавирусы являются возбудителями инфекционного бронхита (куры) и синюшной болезни (индюки).
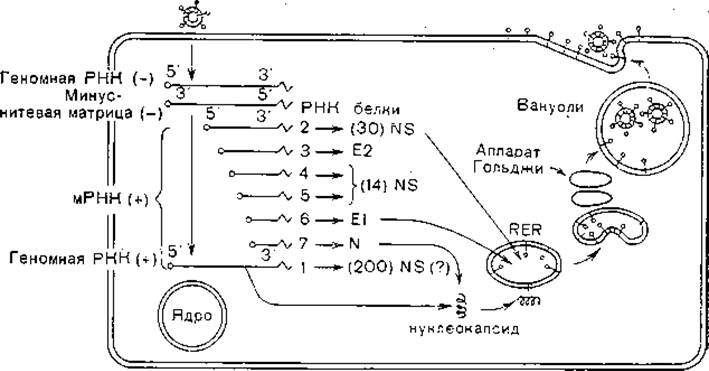
Рис. 19. Репликация коронавирусов (схема). Геномная РНК является мРНК и направляет синтез РНК-пол меразы, которая синтезирует минус-нитевую матрицу. На матрицу минус-нити синтезируются плюс-нитевая геномная РНК и субгеномные мРНК. После трансляции гликопротеиды транспортируются в. аппарат Гольджи, где завершается гликозилирование. Вирионы почкуются во внутриклеточные вакуоли. В скобках — молекулярная масса (х103).
О происхождении коронавирусов сказать что-нибудь определенное трудно. Целесообразно рассматривать их в сравнительном аспекте с другими оболочечными вирусами, имеющими позитивно-полярный геном, — тогавирусами и флавивирусами. У всех этих групп при общности стратегии генома имеются и существенные различия, можно сказать, тактического порядка. У тогавирусов сначала транслируется часть генома, прилегающая к 5'-концу, на которой закодирован полимеразный комплекс. Затем синтезируется субгеномная РНК (через стадию образования минус-нити), которая кодирует синтез структурных белков. Таким образом разобщаются синтез ранних (неструктурных) белков, репликация РНК и синтез поздних (структурных) белков. У флавивирусов, несмотря на непрерывность генома, мРНК синтезируются отдельно с каждого гена, а репликация геномной РНК осуществляется также через стадию синтеза минус-нити. У коронавирусов этот процесс более своеобразен и происходит через синтез минус-нитей как полного генома (репликация РНК), так и субгеномных РНК, которые начинаются с одной и той же точки и различаются по длине, а следовательно, и числу генов. Будем считать эти различия тактическими и тогда можно рассматривать появление трех рассматриваемых семейств как трех тактических вариантов одной и той же стратегии генома.
При рассмотрении эволюции тогавирусов было показано, что сходная стратегия наблюдается у 4 групп вирусов растений (вирус пятнистого вилта томатов, вирусы болезней табака, бромовирусы и вирус мозаики люцерны). Если считать, что все эти вирусы эволюционно связаны между собой, то логично отнести к ним и флавивирусы, и коронавирусы. Однако пока для этого мало оснований и требуются специальные исследования, которые подтвердят или отвергнут такое предположение. Что же касается эволюции коронавирусов, то в этом случае «движущие силы» и пути эволюции были сходны с теми, которые будут описаны в главе 15 парамик- совирусы). Вероятно, основной экологической нишей для коронавирусов явились одомашненные животные и домовые грызуны. От этих вирусов могли «ответвиться» коронавирусы, поражающие человека. Мы слишком мало знаем об этих вирусах и поэтому не можем более конкретно рассматривать возникновение антропонозов.
Здесь же уместно упомянуть о выделении от лошадей в Берне (Швейцария) оболочечного вируса диаметром 120— 140 нм со спиральным типом симметрии нуклеокапсида. В зараженных клетках происходит синтез двух главных (22 000, 20 000) и 4 других (200 000, 80 000—120 000, 32 000, 17000) белков. Несмотря на некоторое сходство с коронавирусами, М. Horzinek и соавт. (1984) предлагают выделить его в отдельное семейство. Геномом его является РНК, по-видимому, позитивно-полярная. Продолговатый нуклеоид представляет собой открытое кольцо со спиральной симметрией. Белок нуклеокапсида имеет молекулярную массу 20 000. Внешняя оболочка содержит пепломеры. Вирус не имеет серологического родства с коронавирусами, но серологически родствен вирусу Бреда, выделенному при энтерите телят, и сходному вирусу Лион-4, а также вирусу, обнаруженному при гастроэнтерите человека. Предложено выделить их в отдельное семейство Toroviridae.