Эволюция вирусов - Жданов В. М. 1990
Специальная часть
Паповавирусы
Паповавирусы составляют компактную группу не очень многочисленных вирусов, образующую семейство Papovaviridae. В него входят два довольно четко очерченных рода — Papillomavirus и Polyomavirus.
Геном представляет собой двунитевую циркулярную ДНК с небольшой молекулярной массой (3х106—5х106). Вирионы имеют форму икосаэдров, их диаметр 45—55 нм, состоят из 72 капсомеров. Иногда встречаются филаментозные формы. ДНК упакована в полости нуклеокапсидов вместе с 2—3 клеточными гистонами. Размножение происходит в ядре. Большинство вирусов обладает онкогенными свойствами [Альт-штейн А. Д., 1982; Matthews R., 1982].
Поскольку эта группа вирусов детально изучена, мы приведем преимущественно данные, необходимые для понимания возможных путей эволюции этой группы вирусов. Следует учесть, что наиболее подробно изучены вирусы полиомы и SV40 и в значительно меньшей степени — представители рода Papillomavirus. Первичная последовательность нуклеотидов расшифрована для нескольких видов, относящихся преимущественно к этому роду. Так, ДНК SV40 содержит 5243 пар оснований и представляет собой двунитевую кольцевую сверх спирализованную структуру (20—24 витка на молекулу). Она является своеобразной минихромосомой, ассоциированной с клеточными гистонами F3, F2 и F2a1. В нейтральной среде сверхспирализованная ДНК имеет коэффициент седиментации :205, в щелочной среде в условиях плавления, при сохранении непрерывности колец этот коэффициент увеличивается до 53S. При разрыве единой нити сверхспирализация исчезает и коэффициент седиментации снижается до 16S, а при разрыве обеих нитей образуется линейная молекула с коэффициентом седиментации 145.
В составе вирионов SV40 имеются 3 белка: VP1 (молекулярная масса 45000—48000), VP2 (32000—35000) и VP3 (23 000). VP1 является главным белком вирионов и составляет их капсид. Капсид состоит из 60 гексонов (построенных из VP1) и 12 угловых пептонов (построенных из VP2 и VP3). Кроме того, в ходе репродукции вируса синтезируются большой (Т) и малый (t) ранние антигены, антиген U, индуцируется СТОА (рис. 41).
На геноме вируса SV40 расположены 3 гена: А, ВС и D (рис. 42). Ген А кодирует антигены Т, t и U, ген ВС— VP1, ген D — VP2 и VP3. Экономичность использования ограниченного по величине генетического материала выражается в том, что, во-первых, используются обе нити ДНК, во-вторых, один и тот же ген кодирует синтез двух разных белков, антигенов Т и t, причем мРНК для синтеза меньшего белка образуется путем сплайсинга. Антиген Т выполняет две функции — он необходим для репликации вирусной ДНК и участвует в трансформации клетки.
Взаимодействие SV40 с чувствительными к нему клетками может быть двух типов — репликативное и интегративное. При репликации вслед за адсорбцией, проникновением вирионов в клетку и частичной депротеинизацией происходит транскрипция генов для ранних белков с помощью клеточной полимеразы и трансляция. При этом образуется антиген Т, инициирующий репликацию вирусной ДНК. Последний процесс происходит при участии клеточного ДНК-полимеразного комплекса. Кроме того, в ранней стадии синтезируются еще 4 белка: средний антиген Т, малый антиген t, антиген U и СТОА. Репликация ДНК начинается в определенном месте (origin of replication). Затем происходят транскрипция поздних (структурных) белков, самосборка вирионов и выход их из деградировавших клеток.
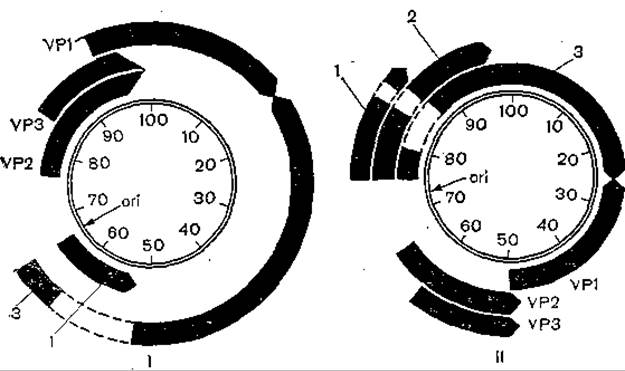
Рис. 41. Карта локализации антигенов и структурных белков SV40 (I) и вируса полиомы (II).
ori — начало репликации ДНК; вирусный геном условно разделен на 100 единиц; Т-антигены: 1 — малый, 2— средний, 3 — большой.

Рис. 42. Физическая, транскрипционная и трансляционная карта геномов вируса полиомы (I) и SV40.
Т — антигены; st — малый, mt — средний, Г — большой.
При интегративном взаимодействии ДНК SV40 встраивается ковалентно в хромосомы зараженных клеток и инфекция приобретает абортивный характер: синтезируются ранние белки, из которых по крайней мере один (антиген Т) обеспечивает трансформацию клетки. Необходимо, однако, отметить, что такое подразделение относительно, так как интеграция может иметь место и при продуктивной инфекции, а интеграционное взаимодействие не исключает развития продуктивной инфекции (синтеза вирионов).
Данные, полученные при изучении SV40 и вируса полиомы, в общем справедливы для всех паповавирусов, однако со значительными коррективами. Так, среди полиомавирусов известны вирусы человека (BK, JC), которые вызывают не неопластические процессы, а демиелинизирующие процессы в мозге. При совместном размножении аденовируса и SV40 образуются гибриды, содержащие в составе аденовируса полный или частичный геном SV40. Этот феномен не наблюдается у других паповавирусов. Вирус бычьей папилломы может перси- стировать в зараженных клетках в виде плазмиды [Roesl F. et al., 1986].
Род Papillomavirus включает в себя вирусы папиллом (доброкачественных и злокачественных) кроликов, коров (5 типов), оленей, собак, лошадей, овец, крыс, человека (31 тип). Молекулярная масса ДНК 5х106, диаметр вирионов 55 нм. Все они имеют общие видоспецифические и (некоторые из них) типоспецифические антигены, не культивируются в клеточных системах. Строение генома одного из полиомавирусов человека, JC, расшифровано [Frisque R. et al., 1984]. Он состоит из 5130 пар нуклеотидов, на нем закодированы 6 белков: ранний антиген Т (t), капсидные белки VP1, VP2 и VP3.
Гомология этого вируса с SV40 составляет 69%, с ВК — 75%. Это свидетельствует об эволюционной общности 3 сравниваемых вирусов.
Вирус папилломы человека реплицируется в кератоцитах в виде эписомы, 50—200 копий ДНК на клетку. При этом не происходит ни интеграция, ни продуктивная репликация [La Porta R., Taichman L., 1982].
Род Poliomavirus включает в себя вирусы, поражающие мышей (полиомы, вирус К), кроликов (RRV), хомяков (HOPV), обезьян разных видов (SV40, STМV, LPV, SA12) и человека (ВК, JC). Эти вирусы обладают выраженными онкогенными потенциями, но нередко вызывают бессимптомную инфекцию. Вирусы человека неонкогенны; вирус JC вызывает демиелинизирующий процесс — прогрессирующую множественно-очаговую лейкоэнцефалопатию [Padgett В. et al., 1971], а вирус ВК обнаружен у лиц с пересаженными органами. Вирусы папиллом птиц имеют сходную организацию с вирусами папиллом млекопитающих [Moreno-Lopez J. et al., 1984].
Считается, что малый антиген поддерживает состояние трансформации, обусловленное большим антигеном [Bikel I. et ah, 1986]. При этом в процесс трансформации включается клеточный ген для белка pp60c-src. Вирусы папилломы, по-видимому, содержат не один, а два или больше трансформированных генов [Yang Y. et ah, 1985]. Имеются веские доказательства в пользу вовлечения вирусов папилломы человека в злокачественный процесс в шейке матки [Duerst М. et ah, 1983]. Вообще вирусы папилломы человека, которых уже насчитывается 31 серовар [Lorinez А., 1986], вызывают разные типы опухолей (папилломы, злокачественные), причем определенные патологические формы «связаны» с разными серо- варами вируса.
Эволюция папилломавирусов шла разными путями: завоевание новых «хозяев», формирование разных сероваров в организме одного и того же «хозяина», смена тканевого тропизма. Первый путь привел к возникновению папилломавирусов человека, коровы, лошади, оленя, овцы, козы, собаки, крысы и других животных, многие из которых были одомашнены. Как правило, видовая специализация сопровождалась потерей патогенности для других видов «хозяев». Одновременно формировалась органно-тканевая специализация, отчетливо просматривающаяся у папилломавирусов человека и коровы. У коровы вирусы сероваров 1; 2 и 5 вызывают фибропапилломы, а вирусы сероваров 3; 4 и 6 сохранили эпителиотропность. Эти различия обусловили дальнейшую дивергенцию между упомянутыми двумя подгруппами [Coggins L. et ah, 1985]. Попав во внутривидовую экологическую нишу, папилломавирусы стали дивергировать, образовав разные серовары, особенно многочисленные у человека (41) и коров (6).
Паповавирусы обнаружены также у птиц (вирус budgerigar fledgling disease) [Lehn H., Mueller H., 1986]. Описаны также рекомбинанты между SV40 и аденоассоциированным вирусом [Grossman Z. et ah, 1984].
Для одних родов характерны серологические связи, в основе которых лежат консерватизм обоих генов и различия, обусловленные другими генами. Применительно к вирусам папилломы консервативными являются гены E1, Е2 и L1 [Fucks Р. et ah, 1986].
Происхождение паповавирусов, четко отграниченных от других, групп вирусов и имеющих столь же четко отграниченные две подгруппы (роды), члены которых взаимосвязаны между собой, имея общие антигены, является загадкой. Экологической нишей обеих групп паповавирусов служат только млекопитающие, что может свидетельствовать о сравнительно недавнем происхождении всего семейства. Серологические связи внутри родов дают еще больше оснований утверждать о сравнительно недавнем их происхождении, и эволюции, измеряемой немногими миллионами лет. Наличие у приматов и человека сходных вирусов позволяет заключить, что эволюция паповавирусов была сопряжена с эволюцией их «хозяев». При этом имеются явные различия между вирусами двух родов — не только в величине их генома, но в типе специализации. Папилломавирусы вызывают неопластические процессы у своих «хозяев», именно эти поражения обеспечивают горизонтальную эстафетную передачу вирусов. Специализация их выражается еще и в том, что каждый из них поражает лишь один вид.
В этом отношении полиомавирусы имеют более широкий (по крайней мере принципиально) круг «хозяев», вызываемые ими патологические процессы более разнообразные, а у «хозяев» инфекция может протекать бессимптомно. В то же время вирусы этой группы или, вернее, часть из них имеют хорошо организованный механизм трансформации, включая поддержание клеток в трансформированном состоянии. В чем биологический смысл существования трансформирующего клетки аппарата у SV40, если у своих «хозяев» он почти никогда не вызывает злокачественной трансформации? Поражает также удивительная экономия генетического материала и мультифункциональность неструктурных белков, особенно антигенов Т (t). Невольно возникает вопрос не только о том, как могли возникнуть такие совершенные автономные генетические структуры, но и для чего все это понадобилось. Этот вопрос пока остается без ответа. Пожалуй, чуть ли не единственным указанием на возможные источники происхождения этой группы вирусов служит наличие гомологии между белком — продуктом гена ORF4 папилломавируса 8 человека и ядерным антигеном 2 вируса Эпштейна — Барр [Fuchs Р. et al., 1986]. И еще одним указанием на возможное происхождение вирусов этой группы может служить следующее. При исследовании библиотеки геномов клеток человека были обнаружены фрагменты, которые гибридизовались с ДНК SV40 значительно интенсивнее, нежели Аlu-последовательности, гибридизацию которых можно было объяснить обилием в последних и регулирующей области гена, кодирующего антиген Т. S. Conrad и М. Botchan (1982) заключают, что повторы длиной 72 пар нуклеотидов генома SV40 соответствуют предполагаемому «активатору» клеточной ДНК. Мы хотели бы добавить, что эти данные могут свидетельствовать и о клеточном происхождении данной последовательности, и о молекулярной конвергенции.
Что же касается дальнейшей эволюции, то ее понять легче. Несомненно, оба рода имели общего предка или общих предков, которые затем дивергировали, дав начало более специализированной группе папилломавирусов и менее специализированной группе полиомавирусов. Эволюция их могла идти и внутри видов, и путем распространения от одного вида к другому. При этом за десятки миллионов лет (особенно это относится к первому роду) существенной эволюции не произошло, и экологически вирусы как бы растекались по поверхности, завоевывая новых «хозяев», но сохраняя консервативными большую часть имеющихся у них генов. Хронический характер вызываемых ими процессов, длительное персистирование в организме — все это позволяло, с одной стороны, сохраниться и эволюционировать вирусам даже в некоторых популяциях, а с другой стороны, медленно и далеко не полностью заполнять занимаемую экологическую нишу в виде того или иного «хозяина». Этот тип сложившихся взаимоотношений между популяциями «вирусов» и «хозяев» охарактеризован в современной литературе о паповавирусах. Несмотря на интенсивное их изучение, вопросов значительно больше, чем ответов на них, даже предположительных.