Эволюция вирусов - Жданов В. М. 1990
Специальная часть
Вирусы оспы
Семейство вирусов оспы (Poxviridae) подразделяется на два подсемейства — вирусы оспы хордовых (Chordopoxvirinae) и оспы насекомых (Eniomopoxvirinae). В первом подсемействе выделены 6 родов вирусов: Orthopoxvirus (оспы), Parapoxvirus (паравакцины), Avipoxvirus (оспы птиц), Capripoxvirus (оспы овец), Leporipoxvirus (миксомы) и Suipoxvirus (оспы свиней). В составе каждого рода имеется либо один, либо (чаще) несколько вирусов. Во втором подсемействе выделены 3 рода на основе морфологических особенностей строения вирионов, молекулярной массы геномов, круга поражаемых «хозяев»: вирусы А оспы шейкокрылых (Colioptera), вирусы В оспы чешуекрылых (Lepidoptera) и вирусы С оспы двукрылых (Diptera). В отличие от вирусов оспы позвоночных вирусы, поражающие насекомых, серологически неродственны как между родами, так и в пределах одного и того же рода. Поэтому и классификацию их следует считать предварительной. Кроме того, несколько вирусов оспы не классифицированы, среди них вирусы оспы плотоядных, слонов, раккунов, вирусы оспы Тана и Яба, а также контагиозного моллюска [Matthews R., 1982].
Морфология вирионов оспы настолько характерна у представителей обоих подсемейств, что не было смысла подразделять представителей, поражающих столь отдаленных филогенетически «хозяев», на таксономические группы более высокого ранга. Вирионы поксвирусов имеют овальную или кирпичеобразную форму с соотношением длин осей 1,2—1,7. Их размер 140—170х220—450 нм, эта величина приближается к размеру наиболее мелких бактерий (риккетсии, хламидии, микоплазмы). Строение их не укладывается в рамки типов симметрии других вирусов и отличается деталями у представителей разных родов. Общим для всех вирусов данного семейства является наличие сердцевины в виде вогнутой линии, внешней оболочки и боковых тел, как бы сплющивающих сердцевину (нуклеоид).
Нуклеоид содержит ДНК, ассоциированную с белками. Способ ее упаковки неясен, хотя из комплекса ДНК— белок выделены 3 белка с молекулярной массой 33 000, 28 000 и 12 000, последний белок богат аргинином. Напрашивается аналогия с упаковкой ДНК хромосом эукариотов гистоновыми и негистоновыми белками. Нуклеоид окружен внутренними оболочками. Внешние оболочки состоят из липидов и углеводов. Оболочки построены сложно и состоят из слоев трубчатых или глобулярных структур. Разные роды вирусов различаются преимущественно деталями строения внешних оболочек.
Геном вирионов представляет собой линейную двунитевую ДНК с молекулярной массой 85x106—250x106, концы которой соединены ковалентно, образуя терминальные петли и инвертированные тандемные повторы. Молекулярная масса генома не может служить таксономическим признаком, так как у серологически родственных вирусов она колеблется от 85x106(вирус паравакцины) до 200x106 (вирус оспы птиц). Содержание Г+Ц у вирусов оспы позвоночных составляет 5— 7,5% от массы вириона; состав нуклеотидов: А — 29,5%, Ц — 20%, Г — 20,6%, Т — 29,9%. Геном вируса осповакцины содержит всего 240 000 пар нуклеотидов и имеет длину 82 мкм. Необычная прочность ДНК этого вируса и других вирусов данной группы обусловлена ковалентными связями комплементарных нитей на обоих концах молекулы ДНК, примерно на расстоянии 50 нуклеотидов от ее концов [см. Слепушкин А. Н., 1982]. Как было показано при изучении вируса осповакцины, ее ДНК, подобно ДНК многих других вирусов, содержит терминальные инвертированные повторы, которые, как предполагается, играют важную роль в репликации ДНК. У вируса осповакцины эти повторы весьма велики — молекулярная масса около 7х106, или около 10 000 пар нуклеотидов. Они могут быть частично транскрибированы, кодируя синтез ранних мРНК [Wittek R. et al., 1980].
На геноме вирусов оспы закодировано несколько сотен белков (глико- и фосфопротеидов) с молекулярной массой 80 000— 200 000. По данным разных авторов, для различных вирусов число вирусспецифйческих белков варьирует. Ранее указывалось, что вирионы осповакцины содержат более 80 белков, суммарная молекулярная масса которых составляет около 60% массы генома [Fenner F., 1979]. У вируса оспы Яба обнаружено 37 структурных белков с молекулярной массой 10 000—220 000 [Fenger.T., Rouhandch Н., 1976]. Более поздние данные свидетельствуют, что вирус осповакцины имеет 279 белков, в том числе 13 гликопротеидов [Carrasco L., Bravo R., 1985]. Пока идентифицированы лишь немногие вирионные и и невирионные белки. Нейтрализация вируса связана с поверхностным белком (58 000), гемагглютинация — с гликопротеидом (85 000). Другие мажорные белки обозначают символами,2Ь, 4а, 4Ь, 6а, 6b, 8, 11b; два белка (4а, 4b) составляют 50% суммарных белков сердцевины, масса которой в свою очередь составляет около 50% массы вириона. В наружных оболочках вириона найдено 8 белков, а на самой поверхности — 5 белков. В составе вирионов имеется или в процессе репродукции образуется не менее 10 ферментов, в том числе РНК-полимераза (вирионная транскриптаза), ферменты, катализирующие образование и метилирование кэп-структур, которые обеспечивают синтез и процессинг мРНК, протеин- и тимидинкиназа. По-видимому, и синтез вирусной ДНК катализируется в основном вирусспецифическими ферментами [Slabaugh М., Mattews С., 1984].
Из нескольких (около 10) больших антигенов вирионов один перекрестно реагирует со всеми «членами» вирусов оспы позвоночных. В пределах родов серологические связи разных вирусов выражены еще больше. На ранних стадиях репродукции вируса осповакцины синтезируется белок с молекулярной массой 19 000, сходный с фактором эмбрионального роста клеток млекопитающих (о возможном значении и происхождении этого белка см. ниже). В связи со сложными процессами синтеза и созревания вирусных белков и морфогенезом вирионов антигенная структура внутриклеточного и внеклеточного вирусов различна.
Репликация вирусов оспы (рис. 61) происходит в цитоплазме ступенчато, включая первичное раздевание (uncoating I), «синтез ранних мРНК и белков, вторичное раздевание (uncoating II), репликацию ДНК, синтез поздних мРНК и белков, входящих в состав вирионов, их гликозилирование и фосфорилирование, образование предшественников вирионов, созревание их и выход из клетки [Shida Н., 1986].
Хотя имеются данные об участии клеточного ядра в репродукции вирусов оспы [Miningan Н. et al., 1985], но оно не обязательно, так как весь цикл репродукции вирусов оспы можно воспроизвести в безъядерных клетках [Villareal Е. et al., 1984]. Вместе с тем репродукция этих вирусов, если даже абсолютно исключить клеточно-зависимые синтезы белка, тесно связана с клеточным метаболизмом. Об этом свидетельствуют возможность репродукции штаммов вируса оспoвакцины, дефектных по некоторым генам (например, по гену тимидинкиназы), возможность замены этого гена геном герпесвируса, а также наличие в геноме вируса достаточно обширных нереплицирующихся областей. Именно на использовании последних в значительной мере основаны генно-инженерные исследования, в результате которых вирус осповакцины со встроенными в него чужеродными генами становится экспрессирующим их вектором (см. ниже).
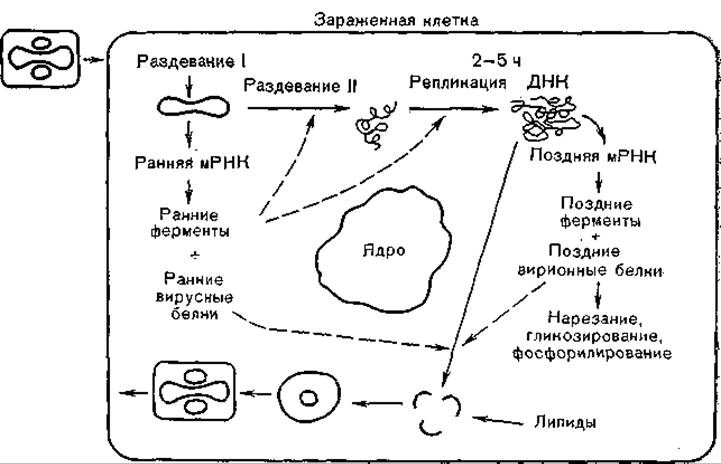
Рис. 61. Цикл репликации вируса осповакцины (схема).
После адсорбции вирионов на клеточных мембранах и проникновения их в клетку вместе с образовавшимися эндоцитозными вакуолями происходят слияние клеточных и вирусных мембран, первичная депротеинизация вирионов и высвобождение сердцевин (нуклеоидов), которые после этой первой стадии раздевания становятся метаболически активными. Синтез ранних мРНК, образование кэп-структур и полиаденилирование обеспечиваются вирионными ферментами, при этом не происходит сплайсинга [Venkatesan S., Moss В., 1981]. Регулирующие области генов вирусов оспы имеют богатые аденином и тимином (ТАТАА) последовательности, но в остальном отличаются от генов про- и эукариотов. В ранних стадиях транскрибируется 14% генома, и из ранних белков синтезируются тимидинкиназа, эндонуклеазы и другие ферменты вируса, а также «родственный белок». Таким образом, депротеинизация вириона включает две стадии: клеточно-зависимую, обеспечивающую удаление внешних оболочек, и вирусзависимую, обеспечивающую более полную депротеинизацию ДНК и возможность ее репликации.
Места, в которых происходят репликация ДНК и формирование вирионов, являются структурами клеточной цитоплазмы, модифицированными вирусом; их обозначают как вирусные фабрики. Более 100 белков, синтезирующихся в первые часы инфекции, обеспечивают вторую стадию раздевания, репликацию вирусной ДНК и транскрипцию поздних генов. Среди ферментов, синтезируемых в этой стадии, следует назвать тимидинкиназу, ДНК-полимеразу, полинуклеотидлигазу, ферменты синтеза ДНК.
Репликация вирусной ДНК связана с образованием разрывов терминальных петель, самопраймированием 3'-концов: вирусной ДНК, но в общем многие детали синтеза вирусной ДНК мало изучены, и поэтому предложено несколько схем ее репликации. Поздняя транскрипция охватывает большую часть генома. На этой стадии транслируются структурные белки, входящие в состав вирионов. Подсчитано, что 7з генома вирусов оспы кодирует ранние, 1/3 — структурные и 1/3 — поздние неструктурные белки [Pennington Т., 1976]. В транскрипции поздних генов, по-видимому, участвует клеточная РНК-полимераза II. Часть белков, синтезируемых на этой стадии, подвергается протеолитическому расщеплению, которое связано, со сборкой вирионов и их предшественников.
Сборка вирионов происходит в «вирусных фабриках» и является многоступенчатым процессом. Сначала дочерняя ДНК ассоциируется с внутренними белками, образуются плотные нити и гранулы. Затем формируется оболочка вирионов в виде гладкой стенки, охватывающей электронно-плотный материал, и образовавшиеся незрелые вирионы (провирионы) выглядят в виде пузырей, в которых формируются нуклеоиды (сердцевины) и боковые тела. Происходят уплотнение внутреннего материала и оболочек и образование поверхностных структур, в результате чего формируются зрелые вирионы. Вирионы выводятся из клетки по каналам внутриклеточного транспорта, а часть их освобождается после гибели зараженных клеток.
Вирусы оспы вызывают глубокое угнетение синтеза клеточных макромолекул, причем эти синтезы блокируются как вирионными белками, так и белками, синтезирующимися в ходе репродукции вируса. Некоторые представители семейства вирусов оспы вызывают пролиферацию зараженных клеток и их: неопластическую трансформацию (вирусы фибромы, Яба, контагиозный вирус моллюска). Для вирусов оспы характерны такие генетические процессы, как рекомбинация внутри родов, спасение маркера [Nakano Е. et al., 1982] и негенетическая реактивация между родами вирусов оспы позвоночных. Вирусы оспы, циркулирующие в природе, подвергаются процессам рекомбинации. Одним из таких рекомбинантов является злокачественный вирус кроликов, явившийся рекомбинантом 10% генома вируса фибромы кроликов и 90% генома вируса миксомы кроликов [Block W. et al., 1985].
Более подробно описание особенностей генетики вирусов оспы можно найти в монографиях и обзорах [см. Гендон Ю. З., 1975]. Здесь же мы хотели бы отметить некоторые важные особенности, характерные, впрочем, не только для вирусов оспы, но и для других, ранее упоминавшихся ДНК-содержащих вирусов (герпесвирусы, бакуловирусы, аденовирусы и др.). Во-первых, ДНК вирусов оспы, близких между собой, или мутантов, легко рекомбинируются, образуя как гомологичные, так и гетерологичные рекомбинанты. В последнем случае образующиеся гетеродуплексы могут включать в себя чужеродные гены. Во-вторых, рекомбинация может происходить не только между полными геномами двух вирусов оспы, но и между полным геномом и сравнительно небольшим его фрагментом (4х106), включенными в плазмиду, при котрасфекции. В-третьих, в геноме вирусов оспы имеются большие участки, которые условно обозначают как несущественные для репликации области генома. Они имеют размер около 25 000 пар нуклеотидов, т. е. более 10% всего генома [Smith G., Moss. В., 1983]. По-видимому, этой областью кодируется синтез белков репликации и других, которые могут быть заменены клеточными белками, как например, тимидинкиназа, а может быть, и белки, обеспечивающие вирулентность вируса. Все эти особенности делают вирус осповакцины весьма удобным вектором для включения в него чужеродных генов. Мы не будем здесь цитировать уже многочисленную литературу, посвященную использованию вируса оспованцины для генно-инженерных операций; сошлемся лишь на меморандум экспертов ВОЗ по этому вопросу [WHO, 1985]. Важно подчеркнуть другое: при генно-инженерных операциях, т. е. искусственных манипуляциях, используют естественные существующие генетические взаимосвязи у этой и других групп вирусов.
Кстати, именно эта особенность вирусов оспы послужила препятствием для применения рекомбинантного вируса осповакцины, содержащего экспрессируемый ген протективного гликопротеида вируса бешенства, для иммунизации лисиц. При поедании мяса, смоченного суспензией такого вируса, у лисиц и других хищников развивается осповакцинный стоматит, сопровождающийся выработкой иммунитета одновременно против оеповакцины и бешенства. Однако комитет экспертов ВОЗ решительно возразил против такого проекта, намечавшегося к проведению в Аргентине. Имелась в виду опасность появления в природе рекомбинантов вирусов оспы с повышенной нейровирулентностью, которая вызвана или связана с геном вируса бешенства, введенного в ДНК вируса осповакцины.
Вирусы оспы вызывают разнообразные заболевания, и все же наиболее выраженно они проявляются в виде высыпаний на коже и слизистых оболочках. Соответственно у животных заболевания передаются при контакте, через корм и питье, а у человека также воздушно-капельным путем. Особое место занимают неопластические процессы, поражающие либо наружные покровы, либо более глубоко лежащие ткани (фиброма). В последнем случае возможно участие в передаче кровососущих насекомых.
Громадная величина генома, сопоставимая с величиной генома у наиболее мелких 'бактерий (микоплазмы, риккетсии и особенно хламидии), автономность процессов репликации и транскрипции, обеспечиваемая вирусспецифическими ферментами, отсутствие сплайсинга — все это позволяет поставить одновременно два вопроса. Закономерно ли считать вирусы оспы вирусами? Не являются ли они продуктами дегенеративной приспособительной эволюции бактерий, причем даже менее далекими, нежели органеллы типа пластид и митохондрий? В самом деле, наиболее мелкие бактерии — хламидии — настолько деградировали эволюционно, что утратили собственные энергорегенерирующие системы и стали своеобразными энергетическими паразитами. По биологическим свойствам от вирусов оспы они-отличаются «только» наличием рибосомных белоксинтезирующих систем. Однако это «только» и является разделом между вирусами и клеточными формами жизни, поэтому пока нет веских оснований исключать вирусы оспы из царства вирусов.
К сожалению, еще не сделана проверка эволюционных соотношений вируса оспы с прокариотами (эукариотами и/или архебактериями), так как все эти проверки, построение филогенетических древ и т. п. делались чаще всего на основе анализа рибосомных РНК (265, 165, 55). Последние имеются даже у потомков далеко эволюционировавших эндосимбионтов (митохондрии, пластиды). А этого как раз и нет у оспенных вирусов.
Кроме того, несмотря на большую автономию генома вирусов оспы от генома клеток «хозяев»-эукариотов, включая процессы репликации вирусной ДНК, имеются факты, свидетельствующие о генетических взаимодействиях между вирусами оспы и их «хозяевами». Так, при исследовании белка с молекулярной массой 19 000, синтезируемого на ранних стадиях репродукции вируса осповакцины, было показано необычное положение цистеиновых и глициновых остатков в молекуле этого белка, сходное с положением этих остатков в двух клеточных белках — трансформирующем факторе роста (TGF) и факторе эпидермального роста (EGF). Эти два белка, обладающие сходными функциями и близкие по величине (50 и 53 аминокислотных остатков соответственно), стимулируют мутагенез. Первый обнаружен во многих опухолях и в эмбриональных тканях, второй — в подчелюстной железе и моче людей. Было высказано предположение, что все 3 белка и некоторые другие белки животных [Blomquist М. et al., 1984] имеют общее происхождение, было даже построено их эволюционное древо (рис. 62). Указанные 3 белка не имеют иммунологического родства, но обладают сходной функциональной активноотью. Белок вируса осповакцины, обладающий свойствами факторов роста клеток, является ранним секретируемым (неструктурным) белком, функции которого пока неизвестны.

Рис. 62. Эволюционное древо белка вируса осповакцины сходных доменов у млекопитающих. Обведенные пунктиром овалы обозначают неопределенность ветвей древа.
С геномами эукариотических клеток геномы оспенных вирусов сближает наличие тандемных повторов в инвертированных терминальных повторах, что было показано при изучении вируса осповакцины. Предполагают, что они облегчают циклизацию однонитевых ДНК во время репликации. Эти повторы имеют размеры 70—150 пар нуклеотидов и повторяются тандемно 13—30 раз [Wittek R., Moss В., 1980].
Таким образом, происхождение вирусов оспы остается неясным. С бактериями его сближают громадный геном, независимая от клетки репликация и отсутствие сплайсинга. Однако структура их генома сходна и со структурой генома эукариотов, а некоторые гены имеют явно эукариотическое происхождение, если не считать их результатом далеко зашедшей молекулярной конвергенции.
Несмотря на большое сходство плана строения и морфологии вирионов оспы позвоночных и оспы насекомых, наличие по крайней мере 4 ферментов с одинаковыми функциями, размножение в цитоплазме и ряд других свойств, указывающих на общность происхождения обеих групп вирусов оспы, в эволюции всей группы вирусов остается много неясного. По-видимому, более древнее происхождение имеют вирусы оспы насекомых уж по одному тому, что насекомые — гораздо более древние формы организмов, нежели теплокровные позвоночные. Вирусы оспы не обнаружены у морских беспозвоночных, и если это факт, а не недостаток знаний, то можно предположить, что вирусы оспы появились не ранее силура, т. е. около 400 млн лет назад, или еще позже, в карбоне.
Освоение ими новой экологической ниши можно связать с развитием кровососущих насекомых, так как некоторые вирусы оспы позвоночных до сих пор могут передаваться кровососущими насекомыми. Кстати, все до сих пор известные вирусы оспы позвоночных поражают теплокровных — млекопитающих или птиц, а вирусы насекомых — как некровососущих (чешуекрылые), так и кровососущих (двукрылые), в том числе комаров-кровососов теплокровных животных. Поэтому образование вирусов оспы позвоночных следует отнести к сравнительно позднему периоду — юрскому или меловому, т. е. 120—150 млн лет назад или еще позже. Именно этим молено объяснить умеренную дивергенцию, существующую между 6 группами (родами) вирусов оспы позвоночных, вплоть до сохранения общих антигенов и еще более широких антигенных связей между вирусами одного и того же рода.
Эволюция этих вирусов происходила путем заполнения ими новых экологических ниш. Она сопровождалась затем, узкой специализацией вплоть до поражения соответствующим вирусом только одного вида. Так появились и эволюционировали вирусы оспы буйволов, верблюдов, кроликов, мышей (группа вирусов осповакцины), узелков доильщиц, контагиозной эктимы, пустулярного стоматита коров (группа вирусов, паравакцины), оспы канареек, голубей, перепелок, ласточек, воробьев, индюшек (группа вирусов оспы птиц), оспы овец, коз, фибромы и миксомы кроликов, оспы свиней. Приручение человеком животных, с одной стороны, усилило специализацию вирусов (оспа овец, коз, верблюдов), с другой — интенсифицировало распространение вирусов, в результате чего одни и те же виды домашних животных стали поражаться разными вирусами оспы. Например, коровы поражаются вирусами осповащины, коровьей оспы, паравакцины, узелков доильщиц; кролики — вирусами кроличьей оспы, фибромы и миксомы. Впрочем, вопрос об узкой специализации не столь уж прост. Мы говорим об оспе коров, но на самом деле речь идет обоспе, которой болеют коровы, но резервуаром которой являются, вероятно, грызуны. Вирус оспы обезьян выделен от белок [Khodakevich L., 1985]. При анализе вируса оспы коров была установлена широкая шкала патогенности, включающая 9 порядков млеко, питающих (рогатый скот, кенгуру, носороги, дельфины, кошачьи, грызуны). Все эти данные позволяют заключить, что грызуны являются основным резервуаром возбудителя в природе и источником заражения коров и других млекопитающих [Berkovitz Е., Pogo В., 1984]. По данным ВОЗ, оспа коров на самом деле является оспой у коров, а оспа обезьян — оспой у обезьян. В последнем случае установлено, что резервуаром оспы обезьян являются белки, а обезьяны — вторичные источники инфекции [WHO, 1986].
При молекулярно-биологическом изучении вируса фибромы Шоупа было показано, что этот вирус наиболее близок к ортопоксвирусам и в то же время имеет часть генома, близкую к геному лепорипоксвирусов [Berkovitz Е., Pogo В., 1985]. Таким образом, рекомбинации вирусов оспы имеют место в природе и некоторые вирусы имеют, несомненно, рекомбинантное происхождение [Berkovitz Е., Pogo В., 1985].
Прежде чем закончить этот раздел, мы хотели бы напомнить об эволюции одного из вирусов оспы — вируса миксоматоза кроликов. Эта эволюция произошла на наших глазах [Fеnner F., 1979]. Данный высоколетальный вирус был использован в Австралии для уничтожения расплодившихся кроликов, вывезенных из Европы и наносивших серьезный урон сельскому хозяйству. Взаимодействие двух популяций — вируса и «хозяина» — привело к развитию персистентной инфекции у животных вместо острой летальной инфекции. Здесь был явно выражен естественный отбор генетически резистентных животных и персистирующего вируса, в различной степени изменивший первоначальный характер инфекции.
Но вернемся к исходным и последующим этапам эволюции вирусов оспы животных [Жданов В. М., Львов Д. К., 1984].
При оспенных заболеваниях птиц и млекопитающих поражаются кожа и слизистые оболочки. Заболевание сопровождается вирусемией и длительным носительством. В этом отношении характерна эктромелия мышей, при которой носительство может быть пожизненным. Все эти заболевания передаются либо при контакте, либо алиментарным путем, либо посредством кровососущих насекомых. Заболевание обычно поражает молодых животных, у которых инфекция может протекать остро. Выжившие животные становятся длительными, нередко, как это имеет место при эктромелии, пожизненными носителями и заражают затем свое потомство.
Инфекции этого типа могли образоваться давно, так как длительность хранения возбудителя и, следовательно, большая продолжительность заразного периода обеспечивали возможность циркуляции вируса в условиях вялотекущих эпизоотий. Тот факт, что к настоящему времени каждый из вирусов оспы поражает один вид или несколько родственных видов, свидетельствует о длительной сопряженной эволюции паразитов с их хозяевами. В этом направлении эволюция вирусов, оспы продолжалась и после одомашнивания животных, а в результате того, что вирусы происходили от нескольких предков, сложилась такая ситуация, что некоторые виды прирученных человеком животных поражаются несколькими вирусами. Как уже упоминалось, у коров наблюдаются коровья оспа, нодулярный дерматит, папулезный стоматит, заболевания, связанные с параоспенными вирусами (контагиозная эктима, «узелки доильщиц» и др.). Все эти заболевания вызываются различными вирусами, отнесенными к разным родам. Кроме того, существует еще одно заболевание — оспа буйволов, вызываемая особым вирусом. Овцы болеют овечьей оспой и пустулярным дерматитом, вызываемыми разными вирусами оспы. Это относится и к некоторым диким животным: кролики болеют кроличьей оспой, фибромой и миксомой. Некоторые из этих заболеваний имеют ограниченные ареалы, но в целом оспенные заболевания домашних животных убиквитарны.
Сравнение оспенных заболеваний диких и домашних животных показывает наличие сходства и различий между ними. Для первых характерны длительный заразный период, наличие носительства и нередко хроническое течение. Из инфекций домашних животных наиболее приближаются к такому типу инфекций заболевания, передаваемые параоспенными вирусами, и, вероятно, осподифтерит кур. В противоположность атому оспа овец, коз, коров, лошадей, свиней и верблюдов характеризуется более острым течением и отсутствием длительного носительства. Такого типа инфекции не могли «укорениться» среди диких животных, в связи с чем можно считать, что оспенные заболевания домашних животных возникли сравнительно поздно, после приручения животных человеком, т. е. в то время, когда уже имелись большие стада этих животных.
Приручение домашних животных и появление больших стад привели к тому, что болезни типа тех, которые вызывают параоспенные вирусы, эволюционировали по пути становления инфекций с более острым течением, что обеспечивало более быстрое распространение возбудителей в стаде. Более острое течение болезни стимулировало выработку более совершенного иммунитета, исключавшего возможность длительной латентной инфекции. Такого типа инфекция не могла бы существовать среди диких животных, но в больших стадах домашних животных при выраженном контакте между ними существование острой инфекции без длительного носительства было возможным. При наличии многих видов животных, прирученных человеком, создавалась возможность взаимного обмена паразитами (вирусами), образования экологических разновидностей, а также самостоятельных заболеваний в результате адаптации вирусов оспы к организму домашних животных. Следует подчеркнуть, что различные оспенные заболевания могли возникнуть среди стадных животных и менее вероятным являлось приспособление вирусов к тем видам животных, которые не разводились в стадах. Не удивительно потому, что известные до сих пор оспенные заболевания типа выраженной острой инфекции не поражают таких домашних животных, как кошки и собаки. Они распространены только среди стадных животных, домашней птицы (осподифтерит кур), а также среди мышевидных грызунов (эктромелия).
Итак, уже на ранних стадиях развития общества (поздний период варварства), когда появились большие стада домашних животных, создались условия для возникновения оспенных заболеваний этих животных с острым и подострым течением. Заражение животных происходило преимущественно алиментарным путем, в связи с чем вирусы вызывают поражение не только кожи, но и слизистых оболочек. Поскольку заражение происходило алиментарным путем, вирусы могли сохраниться лишь при условии их высокой устойчивости к неблагоприятным влияниям факторов внешней среды. И действительно, возбудители оспенных заболеваний диких животных отличаются устойчивостью и способностью длительно сохраняться во внешней среде.
Оспенные заболевания возникли в странах Старого Света, так как в Америке до открытия ее европейцами, кроме лам, не было других видов прирученных животных, а ламы не болеют естественными оспенными заболеваниями. Впервые большие стада домашних животных появились в Юго-Западной и Средней Азии, а также в Северной Африке (Египет). Здесь, по всей вероятности, и возникли оспенные заболевания домашних животных.
Происхождение оспы человека в настоящее время представляется более понятным, чем 20 лет назад. Естественным было бы искать предшественников вируса оспы человека среди вирусов рода Orthopoxvirus, в котором находятся вирусы оспы обезьян и коров, патогенные для человека. Рассматривая этот вопрос ранее [Жданов В. М., 1963], мы отдавали предпочтение вирусу оспы коров. Однако в последние годы в связи с ликвидацией оспы во всем мире внимание исследователей было привлечено к оспенным заболеваниям человека зоонозной природы, в частности к заболеваниям человека оспой обезьян. К началу 1984 г. в некоторых странах Экваториальной Африки было зарегистрировано более 60 таких случаев с несколькими летальными исходами. Лабораторные методы исследования позволяют разграничивать вирусы оспы обезьян и человека, оспы и осповакцины, в частности, выявлять антигены, специфичные для трех наиболее близких вирусов этой группы: МО (антиген вируса оспы обезьян), VА (антиген, общий для вирусов оспы человека и осповакцины, VC (антиген вируса осповакцины). При клонировании штаммов вируса оспы, изолированных от обезьян, С. С. Маренникова и соавт. (1972, 1976) выделили так называемые белые варианты, приближающиеся по своим свойствам к вирусу оспы человека.
Такие же варианты изолированы и от диких грызунов в тех же районах Экваториальной Африки (табл. 21).
На основании данных табл. 21 можно предположить, что источником происхождения вируса оспы человека могли быть белые варианты вирусов оспы обезьян или грызунов Экваториальной Африки. Известно, что вирус оспы обезьян патогенен для человека, он вызывает генерализованный процесс, но в обычных условиях не передается от человека к человеку. Однако нельзя исключить, что болезнь может передаваться от человека к человеку контактным путем, такой путь передачи заразного начала вполне возможен в примитивных бытовых условиях.
Генерализация процесса резко повышает заразность больного, так как в этом случае наличие сыпи на слизистых оболочках полости рта и зева (энантема) и мацерация их создают новые дополнительные возможности для передачи инфекции воздушно-капельным путем. Заразность больного резко возрастает, становится возможным развитие самостоятельных вспышек среди людей. Надо полагать, что такого рода вспышки возникали не раз, однако в условиях первобытно-общинного строя они не могли широко распространиться и ограничились пределами рода или племени.
Таблица 21. Сравнительные свойства некоторых вирусов оспы хFenner F., 19771
Свойства, показатель |
Вирус оспы человека |
Белые варианты |
Вирус обезьян |
осповакцины |
|
от обезьян |
от грызунов |
||||
Оспины на куриных эмбрионах |
Мелкие, белые |
Мелкие, белые |
Мелкие, белые |
Мелкие, розовые |
Большие, белые |
Максимальная температура роста, °С |
37,5—38,5 |
38,5 |
38,5 |
Неизвестно |
Неизвестно |
Рост на |
|||||
коже кролика |
— |
— |
— |
+ |
+ |
клетках ВК13 |
— |
— |
— |
+ |
+ |
клетках Рек Патогенность |
+ |
+ |
+ |
— |
+ |
для мышей |
Низкая |
Низкая |
Низкая |
Высокая |
Высокая |
куриных эмбрионов |
» |
||||
Гемагглютинация Антиген |
Слабая |
Слабая |
Слабая |
Сильная |
Сильная |
Полипептиды Хозяин |
|||||
человек |
+ |
Неизвестен |
Неизвестен |
+ |
+ |
обезьяна |
+ |
+ |
» |
+ |
+ |
мышь |
— |
Неизвестен |
Неизвестен |
+ |
+ |
другие |
— |
» |
Крысы |
Муравьед |
Корова |
Примечание. + да, — нет.
Увеличение скученности населения и числа связей между людьми, а также образование больших государств создавали возможности для более широкого распространения такого типа инфекции. Отдельные штаммы вируса, характеризовавшиеся более выраженной патогенностью и способностью вызвать генерализованный процесс, в этих условиях могли обусловить большие эпидемии. Можно предполагать, что не одна такая эпидемия возникла в это время, не получая дальнейшего развития и «охватив» более или менее значительное число людей, вирус погибал, не имея возможности к дальнейшему распространению в человеческом обществе. Несомненно, что процесс формирования оспенной инфекции человека происходил не только путем отбора мелких изменений вирусов в сторону увеличения их патогенности, усиление патогенности могло произойти и скачкообразно, путем мутации. Один из таких скачков мог привести к резкому повышению патогенности и появлению у вируса способности вызывать генерализованный процесс. В условиях крупного государства, при высокой плотности населения это привело к отрыву возбудителя от прежнего резервуара инфекции и обращению его среди людей. Обширность территории и большое количество, а в ряде случаев и высокая плотность населения обеспечивали непрерывность эпидемического процесса, что было невозможным в условиях первобытнообщинного строя.
Дальнейшая эволюция оспы как инфекции определялась, с одной стороны, условиями развития общества, с другой — характером реакций организма человека. Больше возможностей сохраниться имели те штаммы вируса, которые вызывали острый генерализованный процесс с обильной сыпью, так как такие заболевания сопровождались массивным выделением вируса и заражением большого числа окружающих людей. Поэтому в ходе эволюции отбирались штаммы вируса, вызывающие болезнь с острым течением. Эти штаммы вызывали более мощную реакцию организма с выработкой напряженного иммунитета, что исключало возможность длительного сохранения возбудителя в организме и носительства. Штаммы, вызывавшие слишком тяжелые заболевания с высокой летальностью, также не могли «укорениться», так как вымирание населения приводило к гибели вируса. Таким образом, в результате взаимодействия этих противоречивых влияний формировалась инфекция с острым течением, высокой инфекционностью и стойким постинфекционным иммунитетом, каковой и явилась натуральная оспа.
Остается рассмотреть вопрос о времени и месте образования оспы. Резкая очерченность клинической картины оспы и характерные ее последствия (рубцы) позволяют полагать, что эта болезнь не могла остаться незамеченной даже в древности.
Наиболее древним свидетельством оспы являются находки у египетской мумии IX династии (1200—1100 лет до новой эры) пузырчатой сыпи, которая признается проявлением оспы. Свидетельства в пользу существования оспы в Египте были еще в более древние времена — в папирусах Аменофиса I, относящихся к 3730—3710 гг. до новой эры [Башенин В., 1938]. Вероятно, столь же древней является эта инфекция в Юго-Восточной Азии, так как там еще в XII в. до новой эры были попытки примитивной вакцинации против нее [Вогралик Г., 1935]. F. Burnet (1945) более сдержанно относился к этим данным и считал, что первые достоверные сведения об оспе относятся к VI—XI вв. новой эры.
Приведенные данные позволяют сделать вывод, что местом возникновения оспы явились античные государства Северной Африки, Юго-Восточной Азии и Восточного Средиземноморья, откуда они распространились в другие страны. Оспа возникла на заре цивилизации и поэтому должна считаться одной из наиболее древних инфекций с воздушно-капельным путем передачи.
На примере эволюции оспы мы встречаемся с фактом, имеющим глубокий биологический смысл. Превращение зоонозной инфекции (оспы обезьян) в антропоноз (оспу человека), т. е. отрыв от прежнего резервуара инфекции и адаптация к новому, сопровождалось коренной переменой механизма передачи. Передающаяся контактно-алиментарным путем инфекция животных становится инфекцией человека, передающейся воздушно-капельным путем. Именно эта перемена механизма передачи сделала возможным «укоренение» оспы среди людей, притом лишь на определенной стадии развития человеческого общества. Став эпидемиологически совершенно самостоятельным заболеванием, имеющим две разновидности (оспу и аластрим), современная оспа до сих пор носит на себе «печать» своего происхождения: антигенную близость к оспе обезьян, дерматотропность возбудителя и высокую стойкость вируса. Это мало характерно для инфекций с воздушно-капельным механизмом передачи.
Последующая история оспы прослежена довольно детально. Первая зафиксированная в истории эпидемия оспы имела место в XI в. на Ближнем Востоке во время арабско-абиссинской войны. В VI в. эпидемии возникли в Италии и Франции, а в начале VII в. занесенная арабами в Египет оспа распространилась затем по всему европейскому побережью Средиземного моря. В середине XIII в. эпидемия оспы наблюдалась в Исландии, а в XVI в. опустошительные эпидемии поразили многие страны Европы. С этого времени оспа в Европе стала эндемической болезнью, поражавшей население преимущественно детского возраста. С открытием Америки оспа была завезена на острова Карибското моря и на материк и произвела там опустошительные эпидемии. В XVI в. оспа была почти такой же «обязательной» болезнью, как ныне корь. Г. Вогралик (1935) указывал, что со времени появления оспы в Европе до конца XVIII в. от нее погибло не менее 150 млн человек. По другим данным, в XVII—XVIII вв. в Европе ежегодно болели оспой более 10 млн человек и умирали от нее около 1,5 млн человек. В России в этот период от оспы ежегодно погибали до 0,5 млн человек.
После открытия Э. Дженнером способа иммунизации против оспы на ее истории отражалось активное вмешательство человека в эпидемический процесс. Однако долго еще эта мера оставалась неиспользованной, и только в XX в. в наиболее развитых странах эпидемическое распространение оспы было прекращено, хотя эта болезнь ежегодно продолжала «заноситься» в Европу и Северную Америку главным образам из колониальных стран Юго-Восточной Азии и Африки, даже не предпринимались серьезные попытки ограничить эпидемическое распространение оспы. В Советском Союзе оспа была ликвидирована в 1936 г. и тем самым не только был положен конец ее эпидемическому распространению, но и обеспечена надежная охрана страны от заноса ее из зарубежных стран. Об этом свидетельствует редкость этих заносов в последующие годы, несмотря на большую протяженность границ со странами, высокоэндемичными по оспе, а также значительное расширение экономических и культурных связей со всем миром. Последний занос оспы в СССР (Москву) из Индии был в декабре 1959 г. Не распознанная вовремя вспышка поразила 36 человек, однако была быстро ликвидирована карантинными мерами и иммунизацией всего населения Москвы.
Новый этап в борьбе с оспой наступил в послевоенный период, когда многие страны освободились от колониальной зависимости и стали на путь самостоятельного развития. В этих условиях стала возможной ликвидация оспы во всем мире. По предложению Советского Союза [Жданов В. М. и др., 1959] на Ассамблее ВОЗ в 1958 г. была принята резолюция о ликвидации оспы во всем мире.
В течение первого десятилетия эта работа не приняла должного размаха, и к 1967 г. оспа регистрировалась более чем в 40 странах. Положение изменилось, когда ВОЗ приняла «интенсифицированную программу» искоренения оспы (1967—1976 гг.). Стали проводиться активное выявление больных оспой и иммунизация окружающего больных населения. За выявление больных оспой были назначены денежные вознаграждения. Все эти мероприятия оказались не безрезультатными: сначала число регистрируемых заболеваний увеличилось за счет более волной регистрации, но затем оно стало уменьшаться. «Большая оспа» (Variola major) была побеждена раньше, последний случай был зарегистрирован 16 октября 1975 г. в Бангладеш. Еще через 2 года, 26 октября 1977 г., был зарегистрирован последний случай «малой оспы» (Variola minor) в Сомали. Таким образом, было осуществлено искоренение оспы во всем мире [Ладный И. Д., 1985]. На протяжении этого периода Советский Союз обеспечивал материально и кадрами программу глобального искоренения оспы. Многие советские специалисты работали в странах Азии и Африки вплоть до ликвидации последних очагов оспы, а правительство СССР передало ВОЗ и отдельным странам более 1,5 млрд доз противооспенной вакцины. С 1977 г. оспа не угрожает человечеству, и возбудитель этой болезни сохранен лишь в немногих лабораториях, где продолжается его изучение.