Зоология позвоночных - В. М. Константинов 2011
Хордовые
Наземные, или четвероногие, позвоночные - Позвоночные с зародышевыми оболочками
Класс Пресмыкающиеся, или рептилии
Общая характеристика
Пресмыкающиеся — первые типичные наземные позвоночные животные, все основные черты Amniota у них ярко выражены.
Они дышат только легкими и имеют оформленные дыхательные проводящие пути; механизм дыхания — всасывательного типа при помощи грудной клетки; их тело покрыто кожей с ороговевшими щитками, лишенной желез. У пресмыкающихся прогрессивно изменяются скелет и мускулатура, обеспечивая движение по твердому субстрату и пищевую активность. Особенности размножения характеризуются внутренним оплодотворением, развитием яйцевых и зародышевых оболочек, усилением заботы о потомстве.
Ряд особенностей рептилий свидетельствует о примитивности организации: пойкилотермия, низкий уровень обменных процессов, смешение крови в сердце и др. Общее число видов современных рептилий около 7 000.
Строение пресмыкающихся
(на примере ящерицы рода Lacerta)
Кожные покровы. Кожные покровы рептилий существенно отличаются от кожных покровов амфибий (рис. 92). Верхний слой эпидермиса ороговевает и постоянно слущивается. Его регенерация обеспечивается деятельностью нижнего, живого, слоя эпидермиса. В нем формируются роговые щитки, или чешуи. Ороговение эпидермиса и наличие роговых чешуй имеют важное приспособительное значение, предохраняют тело животных от иссушения. Однако распространенное мнение о непроницаемости кожи рептилий для воды не совсем точно. Мало воды через кожу теряют виды, обитающие в пустынных засушливых областях. У крокодилов же до 75 % всех влагопотерь осуществляется через кожу.
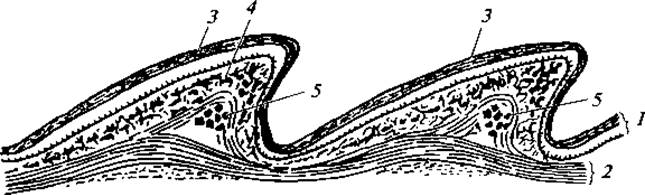
Рис. 92. Продольный разрез через кожу ящерицы (Lacerta):
1 — эпидермис; 2— собственно кожа; 3 — роговой слой; 4 — пигментные клетки; 5 — кожные окостенения
У некоторых видов под роговыми чешуями залегают костные бляшки, развивающиеся как кожные окостенения в кутисе. Кожа рептилий в отличие от амфибий плотно прилегает к телу и не образует столь характерных, как, например, для лягушек, подкожных лимфатических мешков. Кожных желез у рептилий почти нет. У ящериц по внутреннему краю бедер имеется ряд отверстий — бедренных пор, из которых в период размножения выделяется вязкий секрет. Значение бедренных пор невыяснено. Немногочисленные кожные железы развиты у крокодилов, особенно у молодых. Они расположены на спине, на нижней челюсти, в области клоаки. Есть зачатки кожных желез и у змей. Относительно хорошо кожные железы развиты у некоторых черепах.
Скелет. Позвоночник большинства ящериц составлен процельными позвонками. У низших рептилий (агамы, гекконы) тела позвонков амфицельные. Позвоночный столб более расчлененный и подвижный, чем у амфибий. Он состоит из четырех отделов: шейного, грудопоясничного, крестцового и хвостового.
В шейном отделе у ящериц восемь позвонков. Особенностью этого отдела является не только значительно большее, чем у амфибий, число позвонков, но и своеобразное строение (как и у прочих высших позвоночных) двух первых шейных позвонков. Первый шейный позвонок, атлас, или атлант, представляет собой костное кольцо, разделенное связкой на нижнюю и верхнюю половины. Верхнее отверстие служит для соединения головного мозга со спинным; в нижнее заходит зубовидный отросток второго шейного позвонка — эпистрофея. Атлас вращается вокруг этого зубовидного отростка. Данные эмбрионального развития показывают, что зубовидный отросток представляет собой тело первого шейного позвонка, сочленившееся (но у рептилий не срастающееся) с телом эпистрофея. Эта особенность шейного отдела позвоночника обеспечивает подвижность головы.
Грудопоясничный отдел у ящериц состоит из 22 позвонков. Все они несут ребра. Ребра первых пяти позвонков присоединены к грудине и формируют настоящую грудную клетку, свойственную большинству пресмыкающихся (полной грудной клетки нет, например, у змей, у которых нет и грудины). Грудина ящериц хрящевая, эмбрионально она возникает в результате срастания грудных концов ребер.
В крестцовом отделе два позвонка. К их поперечным отросткам причленяются подвздошные кости таза.
Хвостовой отдел включает несколько десятков позвонков. Передние из них несут остистые и поперечные отростки и даже зачаточные ребра. К концу хвоста позвонки утрачивают отростки и приобретают вид палочковидных косточек. Тела почти всех хвостовых позвонков разделены тонкой неокостеневающей прослойкой на передние и задние отделы. При отбрасывании хвоста (аутотомии) разрыв происходит не между двумя позвонками, а посередине какого-либо позвонка, в области этой прослойки. Разрыв обусловливается сокращением специальных мышц хвоста, имеющих вид вставленных друг в друга конусов, вершины которых обращены к корню хвоста.
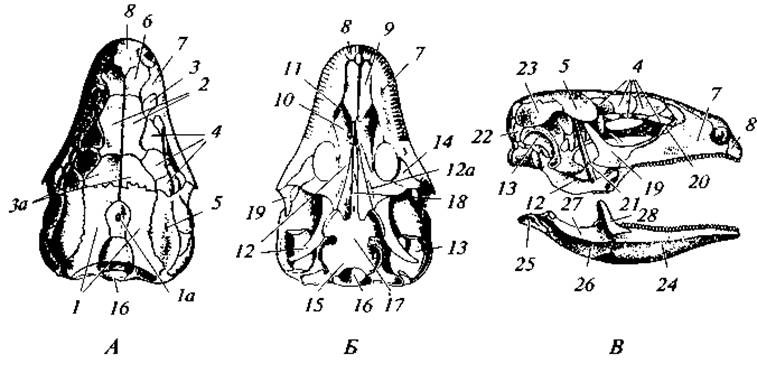
Рис. 93. Череп ящерицы (Lacerta):
вид сверху (А); снизу (Б); сбоку (В). Кости: 1 — теменные (1а — отверстие для теменного глаза); 2 — лобные; 3 — предлобная; 3а — предтеменные; 4 — надглазничные; 5 — заглазничная; 6 — носовая; 7 — верхнечелюстная; 8 — межчелюстные; 9 — сошник; 10 — нёбная; 11 — хоана; 12 — крыловидная (12а — зубы на крыловидной кости); 13 — квадратная; 14 — поперечная; 15 — основная затылочная; 16 — затылочный мыщелок; 17— основная клиновидная; 18 — остаток парасфеноида; 19 — скуловая; 20 — слезная; 21 — столбчатая (надкрыловидная); 22 — чешуйчатая; 23 — надвисочная; 24 — зубная; 25 — сочленовная; 26 — угловая; 27 — надугловая; 28 — венечная
Из всего изложенного видно, что позвоночный столб рептилий характеризуется большей дифференцировкой, обусловливающей лучшую подвижность головы и более прочное прикрепление к осевому скелету пояса задних конечностей. Кроме того, возникновение грудной клетки делает возможным иной, чем у амфибий, механизм воздушного дыхания.
Череп. Особенностью черепа рептилий является почти полное окостенение первичного хрящевого черепа и развитие большого числа кожных костей, формирующих его крышу, бока и дно (рис. 93).
В затылочной области, сформированной четырьмя одноименными костями, характерно развитие только одного (а не двух, как у амфибий) затылочного мыщелка. Область слуховых капсул сложена тремя парами ушных костей. В височной области типично наличие чешуйчатых костей — важного звена в системе прикрепления задней части верхней челюсти к мозговому черепу. Крыша черепа составлена парными носовыми, предлобными, лобными, предтеменными, теменными, группами окологлазничных костей.
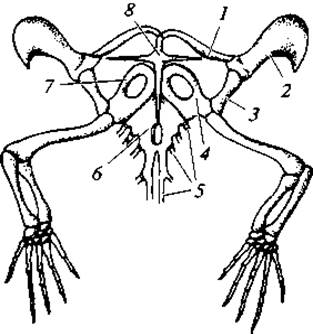
Рис. 94. Плечевой пояс и передние конечности ящерицы (Lacerta):
1 — ключица; 2 — надлопаточный хрящ; 3 — лопатка; 4 — коракоид; 5 — ребра; 6 — грудина; 7 — переднекоракоидный хрящ; 8 — надгрудинник
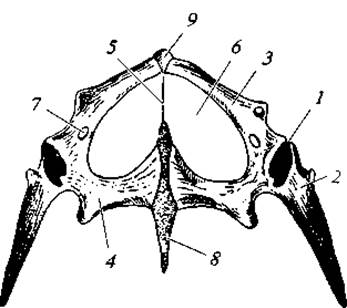
Рис. 95. Тазовый пояс живородящей ящерицы, вид снизу:
1 — суставная впадина для головки бедра; 2 — подвздошная кость; 3 — лобковая кость; 4 — седалищная кость: 5 — связка; 6 — «окно»; 7 — запирательное отверстие; 8 — задний хрящевой отросток; 9 — передний хрящевой отросток
Дно мозгового черепа формируют парные сошники, а также нёбные, крыловидные, квадратные и поперечные кости. Последняя пара сверху прикрепляется к чешуйчатым костям, а снизу с ней сочленяется нижняя челюсть, состоящая из пяти парных костей; зубной, угловой, сочленовной, надугловой и венечной. Верхние челюсти представлены парными и верхнечелюстными костями. Сбоку видна столбчатая кость.
Плечевой и тазовый пояса, а также скелет конечностей не имеют принципиальных отличий от таковых у амфибий (рис. 94, 95).
Мускулатура. Рептилии утратили свойственное низшим позвоночным метамерное расположение мускулатуры. Усиление пятипалых конечностей, появление шейного отдела и большая расчлененность скелета — все это приводит к сложной дифференцировке мышечной системы. Оформляется жевательная, шейная, подкожная мускулатура. Важно появление межреберной мускулатуры, играющей роль в механизме дыхания у всех высших позвоночных.
Органы пищеварения. Органы пищеварения устроены несколько сложнее, чем у амфибий. Это выражается в большей расчлененности пищеварительного тракта и в появлении некоторых новых образований. Ротовая полость заметно отграничена от глотки. У черепах и крокодилов носоглоточные ходы отделены от ротовой полости вторичным костным нёбом. Оно возникает за счет разрастания нёбных отростков межчелюстных и верхнечелюстных костей самих нёбных и крыловидных костей (рис. 96). На дне ротовой полости располагается подвижный мускулистый язык, способный далеко выбрасываться за пределы ротовой полости. Различна форма языка. У змей и многих ящериц он тонкий и часто раздвоенный на конце. У хамелеонов конец языка расширен. Форма языка связана с характером пищи и способами ее добывания.
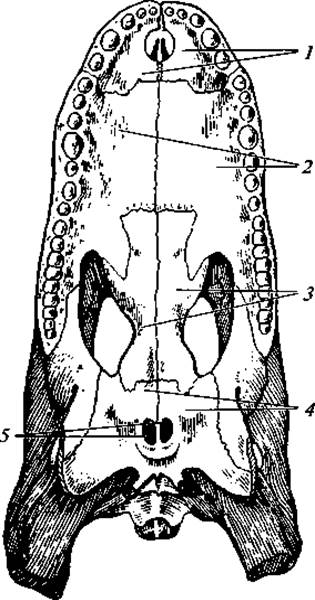
Рис. 96. Вторичное костное нёбо у крокодила:
1 — межчелюстные кости; 2 — верхнечелюстные кости; 3 — нёбные кости; 4 — крыловидные кости; 5 — хоаны
Зубы свойственны большинству рептилий и сидят на верхнечелюстных, межчелюстных, крыловидных и нижнечелюстных костях. На сошнике они есть только у гаттерии. Зубы прирастают к краям соответствующих костей, и только у крокодилов они сидят в альвеолах. У черепах челюсти покрыты роговыми пластинами. Слюнные железы развиты сильнее, чем у амфибий.
Желудок хорошо выражен и снабжен сильной мускулатурой. На границе между тонкой и толстой кишкой находится зачаток кишки, развитой лучше у растительноядных сухопутных черепax. Кишечник заканчивается клоакой. Поджелудочная железа располагается типично, в первой петле кишечника. Печень имеет желчный пузырь, проток которого впадает в кишечник вблизи протока поджелудочной железы.
Органы дыхания. Взрослые рептилии дышат легкими, в связи (развитием рогового покрова кожное дыхание у них отсутствует. Дыхательная система в целом претерпевает ряд изменений. Как у амфибий, общая форма легких мешковидная, однако внутренняя полость значительно уменьшена, так как от стенок легких внутрь отходит сложная сеть перегородок, делящих внутреннюю полость на множество мелких ячей. Особенно сильно развиты они у черепах и крокодилов. У наиболее примитивной современной рептилии — гаттерии — внутренняя полость легких еще велика.
У многих видов ящериц и особенно у хамелеонов задняя часть легких не имеет ячей и перегородок и часто вытянута в виде тонкостенных пальцевидных выростов — легочных мешков. Окисления крови в них не происходит.
Выделение шейного отдела сопровождается дифференцировкой дыхательных путей. Оформляется гортань, поддерживаемая непарным перстневидным и парным черпаловидным хрящами. От гортани отходит длинная трахея, которая делится на два бронха, идущие в легкие.
Механизм дыхания иной, чем у амфибий. Воздух не заглатывается ртом, а втягивается в легкие и выталкивается обратно путем расширения и сужения грудной клетки, обусловленных движением ребер и межреберной мускулатуры. Такой тип дыхания свойствен высшим позвоночным. Более сложная структура легких обеспечивают и более совершенный газообмен.
Увеличивается частота вентиляции легких. Она меняется в зависимости от температуры внешней среды, что имеет некоторое терморегуляционное значение. Так, у некоторых ящериц при температуре воздуха 15°С частота дыхательных движений в минуту — 26, при 25°С — 31, а при 35°С — 37. У некоторых пустынных черепах (например, Testudo sulcata) при значительном повышении температуры воздуха резко возрастает отделение слюны, которая, вытекая изо рта. смачивает нижнюю часть головы, шею и конечности. Испарение слюны с этих поверхностей существенно увеличивает теплоотдачу. Однако эти приспособления имеют ограниченное значение и не могут обеспечить постоянство температуры тела и поддержание ее на высоком уровне.
В эмбриональном состоянии газообмен у рептилий осуществляется с помощью аллантоиса.
Органы кровообращения. Органы кровообращения рептилий в большей степени, чем у амфибий, соответствуют наземному образу жизни и связанному с ним легочному дыханию. Это выражается в первую очередь в более полном разделении артериального и венозного потоков крови, что обусловлено изменениями в сердце, в артериальной и венозной системах.
Сердце у большинства рептилий, как и у амфибий, трехкамерное. Перегородка между предсердиями всегда полная. В желудочке имеется неполная перегородка (рис. 97). Она отходит от брюшной стороны желудочка и в состоянии систолы делит его на короткий момент на левую и правую части. У крокодилов эта перегородка почти полная, и сердце можно считать четырехкамерным.
Сердечный индекс заметно больше, чем у амфибий (до 2,1). Возрастает и частота пульса: у прыткой ящерицы она равна 65 ударам в минуту. Следовательно, кровоток у рептилий более быстрый, чем у амфибий, что является важной предпосылкой интенсификации обмена веществ.
Артериальная система имеет ряд существенных особенностей, oртериальный ствол разделен на три сосуда, самостоятельно отходяших от различных частей желудочка. От правой части желудочка (содержащей венозную кровь) отходит общий легочный сосуд, который вскоре делится на левую и правую легочные артерии. От левой части желудочка (содержащей артериальную кровь) отходит правая дуга аорты, от нее, в свою очередь, — сонные и подключичные артерии. Наконец, от середины желудочка отходит левая дуга аорты. Обогнув сердце, она соединяется с правой дугой аорты и образует спинную аорту.
В связи с такой дифференцировкой артериальных сосудов в легочные артерии поступает только венозная кровь; в правую дугу аорты, а следовательно, и в сонные и подключичные артерии — чистая артериальная кровь. Только в левую дугу аорты поступает смешанная кровь, и, следовательно, в спинной аорте кровь также смешанная, но с явным преобладанием артериальной. Спинная аорта тянется под позвоночником и отсылает ветви к внутренним органам и мускулатуре. В области таза от спинной аорты отходят крупные подвздошные артерии, несущие кровь к задним конечностям и к хвосту.
Венозная система рептилий не имеет столь существенных особенностей, как артериальная (рис. 98). Из хвостового отдела тела кровь собирается в хвостовую вену, которая делится на две тазовые вены. Тазовые вены принимают в себя сосуды от задних конечностей, после чего, отделив две воротные вены почек, они объединяются в брюшную вену. Последняя, приняв ряд вен от внутренних органов, впадает в печень, где образует воротную систему кровообращения.

Рис. 97. Сердце ящерицы:
1 — общий ствол сонных артерий; 2 — внутренняя сонная артерия; 3 — наружная сонная артерия; 4 — легочная артерия; 5 — дуга аорты; 6 — правая дуга аорты; 7 — подключичная артерия; 8 — легочная вена; 9 — полая вена (нижняя) и две вены (верхние); 10 — спинная аорта; 11 — желудочно-кишечная артерия (к внутренностям)
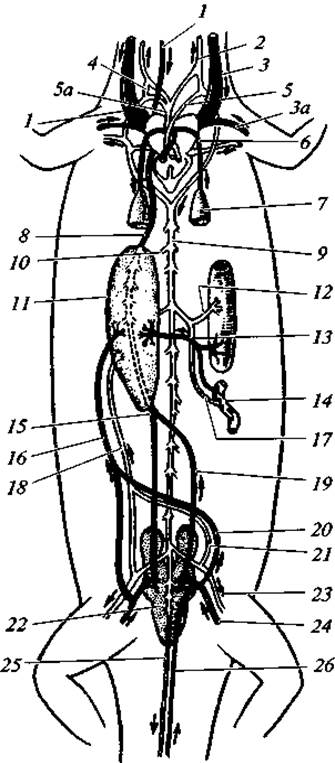
Рис. 98. Схема кровообращения ящерицы:
1 — передняя полая вена; 2 — сонная артерия; 3 — яремная вена; За — подключичная вена; 4 — сонный проток; 5 — левая дуга аорты; 5а — правая дуга аорты; 6 — легочная артерия; 7 - легкое; 8 — нижняя полая вена; 9 -спинная аорта; 10 — межпозвоночная артерия; 11 — печень; 12 — кишечная артерия; 13 — воротная вена печени; 14 — тонкие кишки (отрезок); 15 — задняя полая вена; 16 — брюшная вена 17 — брыжеечная артерия; 18 — наружная брыжеечная артерия; 19 — почечная вена (выносящая); 20 — левая тазовая вена; 21 — левая тазовая артерия; 22 - почка; 23 — артерия задней конечности; 24 — вена задней конечности; 25 — хвостовая артерия; 26 — хвостовая вена
Вены, выносящие кровь из почек, образуют основной венозный сосуд туловищной области — нижнюю полую вену. Она тянется под позвоночником и изливает кровь в правое предсердие. В нес впадает печеночная вена, прошедшая через воротную систему сосудов печени.
От головы кровь собирается в парные яремные вены, которые, соединившись с парными подключичными венами, образуют две (левую и правую) передние полые вены. Они впадают в правое предсердие. В левое предсердие изливают кровь легочные вены. У некоторые видов перед впадением в сердце они объединяются в один сосуд.
Органы выделения. Органы выделения взрослых рептилии представлены тазовыми почками (метанефрос). Они развиваются позади зачатков туловищных почек из общей с ними зачатковой ткани. Туловищные почки формируются как зародышевые органы выделения, функционирующие до вылупления молодых животных из яйца и некоторое время после вылупления. При развитии тазовой почки от вольфова канала отделяется канал, соединяющийся с выделительными трубочками новой почки. Таким образом формируется мочеточник. Левый и правый мочеточники впадают со спинной стороны в клоаку. С брюшной стороны в клоаку открывается мочевой пузырь. У крокодилов, змей и некоторых ящериц мочевой пузырь недоразвит.
После образования тазовых почек туловищные почки у самок практически редуцируются. Однако у самцов передние их части сохраняются и представляют собой придатки семенников, через которые проходят семявыносящие канальцы.
Заметно изменяется структура тазовой почки. В ней возрастает число нефронов: в среднем у рептилий их около 5 тыс. Существенно меняется строение нефронов: достаточно явственно развитые сосудистые клубки есть только у черепах и крокодилов, т. е. у форм, ведущих в той или иной мере полуводный образ жизни. У чешуйчатых сосудистые клубки развиты слабо. Мочеотделение в этом случае осуществляется преимущественно за счет секреции извитыми канальцами нефронов. В соответствии со сказанным скорость фильтрации мочи у рептилий значительно медленнее, чем у амфибий.
Моча у подавляющего большинства рептилий кашицеобразная. Основным продуктом азотистого обмена в аридных условиях служит мочевая кислота — вещество, плохо растворимое в воде, для ее выведения требуется ничтожно малое количество воды. В общеизвестной триаде конечных продуктов азотистого обмена — аммиак- мочевина — мочевая кислота — последняя обладает наименьшей токсичностью. Такой тип обмена сложился в связи с принципиально иными сравнительно с амфибиями условиями эмбрионального развития рептилий, сопровождающегося наличию яйцевых и зародышевых оболочек. В этом случае принято говорить о «замкнутых» яйцах в отличие от «открытых» яиц рыб и амфибий. Важно подчеркнуть, что откладывание мочевой кислоты в системе яйца при развитии зародыша не грозит самоотравлением.
Органы размножения. Половые железы самцов лежат в полости тела по бокам позвоночника. К семенникам примыкают придатки, представляющие, как сказано, остатки мезонефроса. Канальцы придатка впадают в вольфов канал, служащий у самцов рептилий (как и у остальных высших позвоночных) только семяпроводом. Парные семяпроводы выводят мочевую кислоту в клоаку. Все рептилии, кроме гаттерии, имеют совокупительные органы. У ящериц и змей это парные выросты задней стенки клоаки, которые в период возбуждения выворачиваются наружу (рис. 99). У крокодилов и черепах копулятивный орган непарный.
У самок формируются парные гроздевидные яичники. В качестве яйцеводов функционируют мюллеровы каналы, представляющие собой тонкостенные трубки, открывающиеся воронками в полость тела, а другим концом — в клоаку. В среднем отделе яйцевода имеется железа, выделяющая белковую оболочку яйца. В нижней части яйцевода находятся железы, формирующие пергаментообразную, а у черепах и крокодилов — известковую оболочки яйца. Вольфовы каналы у самок не сохраняются.
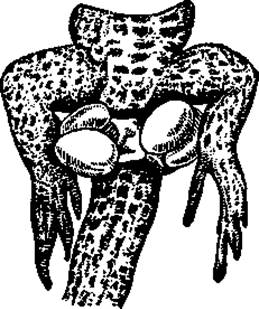
Рис. 99. Наружные выпячивания клоаки самца ящерицы. Видны два выпятившихся копулятивных мешка
Нервная система. Нервная система рептилий более совершенна, чем у амфибий. Полушария переднего мозга относительно крупнее. Большая часть переднего мозга состоит из полосатых тел, кора представлена архипаллиумом. В связи со значительным развитием полушарий переднего мозга промежуточный мозг сверху почти не виден. Заметен эпифиз и хорошо развит теменной орган. Он помешается в отверстии межтеменной кости и весьма эффективно воспринимает световые раздражения (рис. 100). У гаттерий теменной орган своим строением напоминает глаз. В нем различают уплотненный прозрачный передний отдел, в известной мере подобный хрусталику, и бокалообразную заднюю часть, внутренняя стенка которой имеет чувствующие и пигментные клетки — она может быть уподоблена ретине. Средний мозг увеличен.
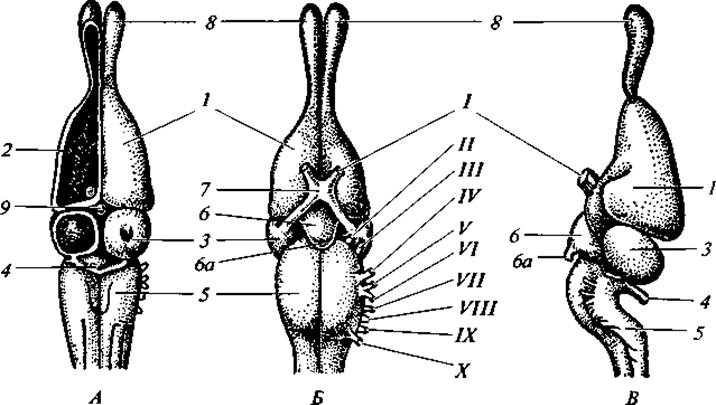
Рис. 100. Головной мозг ящерицы:
вид сверху (А); снизу (Б); сбоку (В); 1 — передний мозг; 2 — полосатое тело 3 — средний мозг; 4 — мозжечок; 5 — продолговатый мозг; 6 — воронка; 6а — гипофиз; 7 — хиазма; 8 — обонятельная доля; 9 — эпифиз; I - Х — черепные нервы
Сильно развит мозжечок. Продолговатый мозг образует в вертикальной плоскости ясный изгиб, что характерно для всех высших позвоночных. Черепных нервов XI пар.
Поведение рептилий по сравнению с амфибиями иное, демонстрирующее возможности более сложных приспособлений при проявлении врожденных рефлексов и инстинктов: у них в 2 раза быстрее формируются новые условные рефлексы.
Органы чувств. Органы чувств пресмыкающихся в значительно большей мере, чем у амфибий, соответствуют наземному образу жизни. Механические раздражения воспринимаются так называемыми осязательными «волосками», расположенными на чешуйках и связанными с осязательными пятнами — скоплениями чувствующих клеток, лежащими под эпидермисом.
Высока чувствительность органов обоняния. Серединная часть носового хода делится на нижний, дыхательный и верхний, собственно обонятельный, отделы. В начале носового хода обособляется преддверие, а его задний отдел, открывающийся в глотку, представляет собой носоглоточный ход (рис. 101).
Есть специализированный якобсонов орган — извитая и слепо заканчивающаяся полость, отходящая вверх от крыши ротовой полости. Считают, что это образование служит для восприятия запахов пищи, уже находящейся во рту. К тому же многие рептилии (например, ящерицы) ощупывают различные предметы при помощи далеко выдвигаемого языка. Втягивая язык в ротовую полость, они переносят мельчайшие частицы предметов в рот, где их запах воспринимается якобсоновым органом.
Орган зрения — глаза снабжены подвижными веками. Нижнее развито сильнее и более подвижно. Имеется третье веко — мигательная перепонка, закрывающая глаз из его переднего угла. У змей и гекконов верхнее и нижнее веки сращены и прозрачны. Аккомодация более совершенная, чем у амфибий. Ресничная мышца имеет поперечнополосатую мускулатуру и не только перемещает хрусталик, но и несколько меняет его форму, что в условиях наземной среды улучшает рассматривание предметов, находящихся на разном расстоянии.
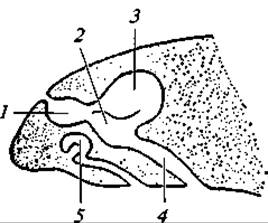
Рис. 101. Обонятельный мешок и якобсонов орган у ящерицы:
1 — преддверие; 2 — дыхательный отдел; 3 — обонятельный отдел; 4 — носоглоточный ход; 5 — якобсонов орган
Орган слуха, как и у амфибий, включает внутреннее и среднее ухо. Среднее содержит одну косточку — стремя. Перепончатый лабиринт более дифференцирован; в нем обособлен мешкообразный выступ — зачаток улитки. У ящериц намечается наружный слуховой проход. Рептилии воспринимают звуки в диапазоне 20 — 6 000 Гц.
У некоторых змей (удавов, гремучих) на щитках верхних и нижних челюстей имеются ямки, иннервируемые ветвями тройничного нерва. Они способны воспринимать тепловые излучения, исходящие от теплокровных животных (потенциальной добычи), на расстоянии 1 — 2 м. Полагают, что эти органы могут различать разницу температуры в 0,001°С.
Систематический обзор современных пресмыкающихся
Сравнительно с амфибиями класс Пресмыкающиеся значительно более многочисленный. Его представители более разнообразны и широко распространены. Общее число современных видов примерно 7 500.
Современные рептилии, составляющие ничтожную часть класса, процветавшего в мезозойскую эру, группируются следующим образом:
Подкласс Анапсидные (Anapsida).
Отряд Черепахи (Testudines, или Chelonia).
Подкласс Лепидозавры (Lepidosauria).
Отряд Клювоголовые (Rhynchocephalia).
Отряд Чешуйчатые (Squamata).
Подкласс Архозавры (Archosauria).
Отряд Крокодилы (Crocodylia).
Рассмотрим некоторые группы.