Биохимические основы жизнедеятельности организма человека - Волков Н.И., Несен Э.Н. 2000
Биохимия спорта
Биоэнергетика мышечной деятельности
Аэробный механизм ресинтеза АТФ
Аэробный механизм ресинтеза АТФ в обычных условиях обеспечивает около 90 % общего количества АТФ, ресинтезируемой в организме. Ферментные системы аэробного обмена расположены в основном в митохондриях мышц. Механизм аэробного окисления питательных веществ носит название «окислительное фосфорилирование», сущность которого описана в главе 3. Общая схема аэробного механизма ресинтеза АТФ в мышцах представлена на рис. 129.
В качестве субстратов аэробного окисления используются глюкоза, высшие жирные кислоты, отдельные аминокислоты, кетоновые тела, молочная кислота и другие недоокисленные продукты метаболизма. Все эти вещества постепенно превращаются в единое вещество — ацетил-КоА, который далее окисляется в цикле лимонной кислоты до конечных продуктов СО2 и Н2О с участием многочисленных окислительных ферментов и вдыхаемого кислорода, доставляемого к тканям гемоглобином эритроцитов крови, а в скелетных мышцах — с участием кислорода, накапливаемого белком миоглобина (см. рис. 129). Энергия окисления накапливается в восстановленной форме переносчиков водорода НАДН2 и ФАДН2, которые передают высокоэнергетические электроны по дыхательной цепи на вдыхаемый кислород, а протоны водорода создают на мембране митохондрий протонный градиент (Н+), который является движущей силой для генерирования АТФ в процессе окислительного фосфорилирования. Когда разница протонного градиента достигает определенной величины (200 мВ), протоны движутся через мембрану митохондрий и взаимодействуют с кислородом с образованием Н2О. Энергия движения Н+ аккумулируется в молекулы АТФ при их синтезе из АДФ и Н3РО4 высокоспецифическим ферментом АТФ-синтетазой.
Скорость образования АТФ в процессе окислительного фосфорилирования зависит от следующих факторов:
✵ соотношения АТФ/АДФ; при отсутствии в клетке АДФ синтез АТФ не происходит;
✵ количества О2 в клетке и эффективности его использования;
✵ активности многочисленных окислительных ферментов;
✵ количества систем дыхательных ферментов в митохондриях;
✵ целостности мембран митохондрий;
✵ количества митохондрий в клетке;
✵ концентрации гормонов, ионов Са2+ и других регуляторов процессов аэробного окисления веществ.
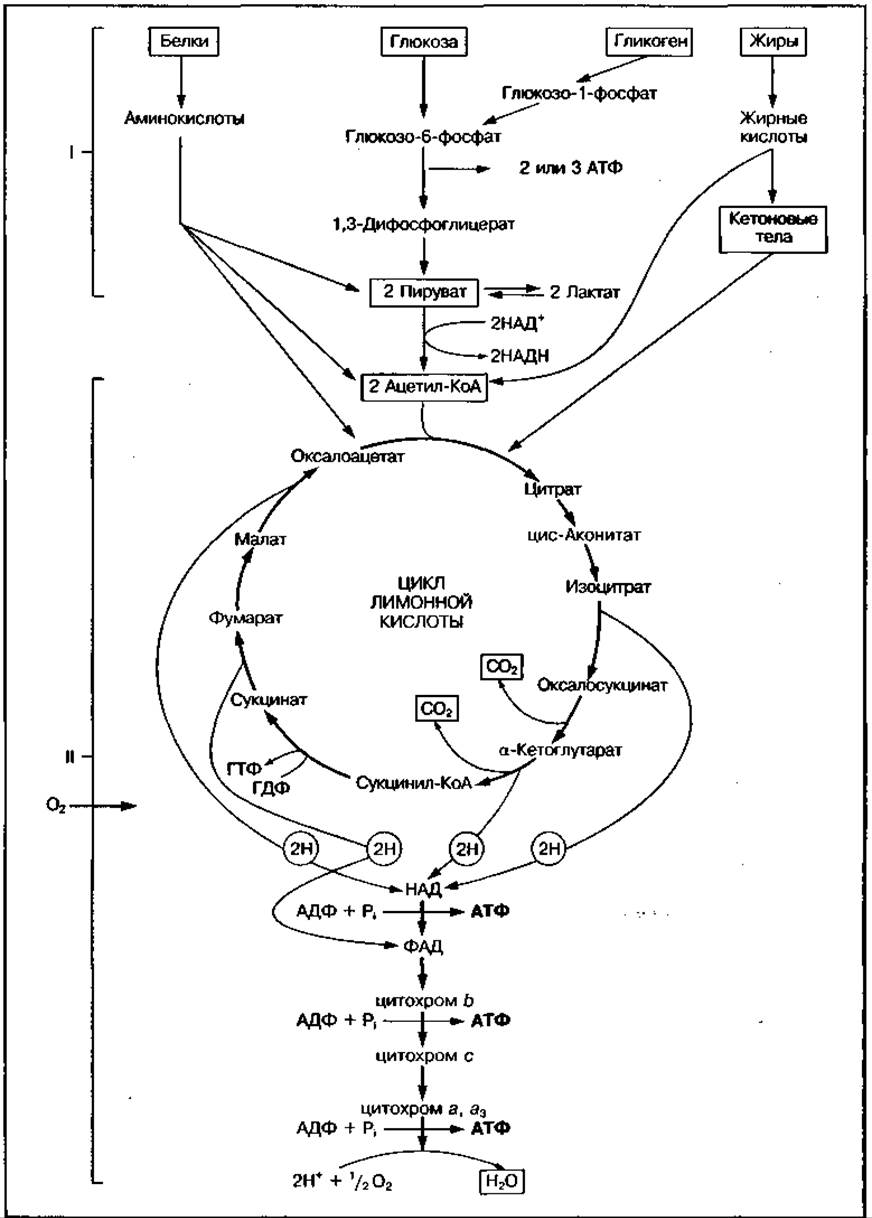
Рис. 129 Схема аэробного механизма ресинтеза АТФ, включающего процессы анаэробного превращения (І) и аэробное окисление — тканевое дыхание (II)
Снижение концентрации АТФ, что наблюдается сразу после начала выполнения интенсивной физической нагрузки, активирует дыхательную и сердечно-сосудистую системы, доставляющие кислород к клеткам.
Количество кислорода, потребляемого легкими, прямо пропорционально количеству О2, используемому в процессах окислительного фосфорилирования. Это позволяет определять величину аэробного энергообразования по поступлению кислорода. Нормализация частоты дыхания и ЧСС происходит только после удовлетворения повышенных потребностей клеток в АТФ.
При потреблении одинакового количества кислорода объем выполненной работы будет большим в том случае, если энергетическим субстратом будут углеводы, а не жиры. Углеводы являются более эффективным "топливом" по сравнению с жирами, так как на их окисление требуется на 12 % меньше кислорода в расчете на молекулу синтезированной АТФ. Поэтому в условиях недопоступления кислорода при физических нагрузках энергообразование происходит в первую очередь за счет окисления углеводов. Поскольку запасы углеводов в организме ограничены, ограничена и возможность их использования в видах спорта, требующих проявления общей выносливости. После исчерпания запасов углеводов к энергообеспечению работы подключаются жиры, запасы которых позволяют выполнять очень длительную работу. Так, в марафонском беге за счет использования мышечного гликогена работа мышц продолжается в течение 80 мин. Часть АТФ может быть получена за счет мобилизации гликогена печени. Следовательно, за счет углеводов можно обеспечить энергией 75 % марафонской дистанции. Остальное количество энергии образуется за счет окисления жирных кислот. Учитывая, что жирные кислоты содержат большое количество энергии, весьма важно развивать способность организма спортсмена к более ранней их мобилизации для энергообеспечения работы. Для этого рекомендуется периодически использовать в тренировке аэробные нагрузки — бег на сверхдлинные дистанции (по 30—40 км и более).
В качестве субстрата окисления могут использоваться и белки, которые распадаются на аминокислоты, способные превращаться в глюкозу или другие метаболиты аэробного процесса окисления. Однако вклад белков в образование энергии при мышечной деятельности составляет всего 5—10 %.
Максимальная мощность аэробного механизма наименьшая и составляет 1,2 кДж ⋅ кг-1⋅ мин-1 и в равной степени зависит от скорости поступления и скорости утилизации О2 в клетках. Мощность аэробного энергообразования оценивается по величине максимального потребления кислорода (МПК), достигнутого при выполнении мышечной работы. У спортсменов эта величина составляет в среднем 5,5—6 л ⋅ мин-1, а у неспортсменов — 2,5— 3,5 л ⋅ мин-1. Поскольку она отражает скорость потребления О2 в работающих мышцах, а на скелетные мышцы приходится большая часть активной массы тела, то в целях сравнения аэробных способностей разных людей величины МПК обычно выражают в расчете на 1 кг массы тела. У молодых людей, не занимающихся спортом, МПК составляет 40—45 мл ⋅ кг-1⋅ мин-1 (800—1000 Дж ⋅ кг-1⋅ мин-1), у спортсменов, занимающихся видами спорта на выносливость, — 80—90 мл ⋅ кг-1⋅ мин-1 (1600—1800 Дж ⋅ кг-1⋅ мин-1).
Максимальная мощность аэробного процесса достигается на 2—3-й минуте неинтенсивной работы у спортсменов и на 4—5-й минуте — у неспортсменов и может поддерживаться до 15—30-й минуты. В более длительных упражнениях она постепенно уменьшается. При марафонском беге средний уровень аэробной энергопродукции составляет 80—85 % максимальной аэробной мощности.
Наиболее интенсивно протекают процессы аэробного энергообразования в медленносокращающихся мышечных волокнах. Следовательно, чем выше процентное содержание таких волокон в мышцах, несущих основную нагрузку при выполнении упражнения, тем больше максимальная аэробная мощность у спортсменов и тем выше физическая работоспособность при продолжительной работе.
Метаболическая емкость аэробного механизма практически безгранична, поскольку имеются большие запасы энергетических источников, дающих большое количество ресинтезируемой АТФ. Так, при окислении 1 молекулы глюкозы в аэробных условиях образуется 38 молекул АТФ, тогда как в анаэробных — только 2 АТФ:
![]()
При окислении высших жирных кислот, например пальмитиновой, образуется еще больше энергии:
![]()
Эффективность энергообразования этого механизма также высокая и составляет около 50 %. Определяется она по порогу анаэробного обмена (ПАНО): у нетренированных людей ПАНО наступает при потреблении кислорода примерно 50 % от уровня VО2max, а у высокотренированных на выносливость — при 80—90 % МПК. Увеличение показателя ПАНО под влиянием специальной тренировки связано с повышением (адаптацией) возможностей кислородтранспортной системы, а также ферментативных, регуляторных и других систем.
Аэробный механизм энергообразования является основным при длительной работе большой и умеренной мощности: беге на дистанции 5000 и 10 000 м, марафонском беге на 25 000 м, велогонках, плавании на 800 и 1500 м, беге на коньках на 5000 и 10 000 м. Он является биохимической основой общей выносливости.