Основы биохимии - А. А. Анисимов 1986
Нуклеиновые кислоты
Биосинтез нуклеотидов
4.5.1. Пути синтеза мононуклеотидов. Пуриновые и пиримидиновые нуклеотиды могут синтезироваться de novo, т. е. из простых предшественников, а также непосредственно из готовых пуриновых и пиримидиновых оснований. Относительное значение этих двух путей существенно отличается для разных клеток. Так, в тканях млекопитающих нуклеотиды преимущественно синтезируются de novo, хотя в быстро растущих тканях образуются обоими путями. Напротив, для нормального роста и развития многих видов бактерий необходимо наличие в питательной среде готовых молекул пуринов или пиримидинов.
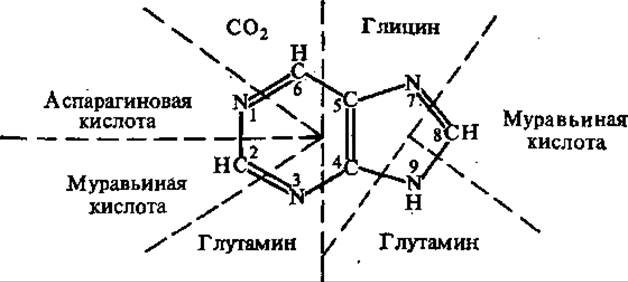
Рис. 4.10. Происхождение атомов пуринового кольца
4.5.2. Биосинтез пуриновых нуклеотидов. Биосинтез пуринового кольца de novo протекает одинаково у разных видов живых существ: у бактерий, дрожжей, птиц, человека. Это один из многих примеров единства ряда основных биохимических процессов всего живого. Эксперименты с использованием изотопов позволили установить, из какого предшественника поступает каждый атом пуринового кольца (рис. 4.10).
Цепочку из двух атомов углерода и одного атома азота (С-4, С-5, N-7) дает аминокислота ели, один атом азота (N-1) поступает из асп, оставшиеся два атома азота (N-3 и N-9) дает глн, два атома углерода (С-2 и С-8) — муравьиная кислота. В первых этапах синтеза участвует D-рибозо-5-фосфат, и уже. над ним надстраивается каркас молекулы гипоксантина. Таким образом, синтезируется не свободное азотистое основание, а его нуклеотид — инозиновая кислота (рис. 4.11).
Инозиновая кислота служит предшественником всех остальных пуриновых нуклеотидов. Так, адениловая кислота (АМФ) образуется в результате аминирования ИМФ, аминогруппа поставляется аспарагиновой кислотой. Образование гуаниловой кислоты (ГМФ) из ИМФ является двухстадийной реакцией. Первая стадия состоит в окислении ИМФ до ксантозин-5'-фосфата (КМФ). Затем происходит его аминирование, и образуется ГМФ. Донор аминогруппы, по-видимому, различен у ряда организмов: у птиц и млекопитающих донором служит глн, в бактериальных системах — NH3.
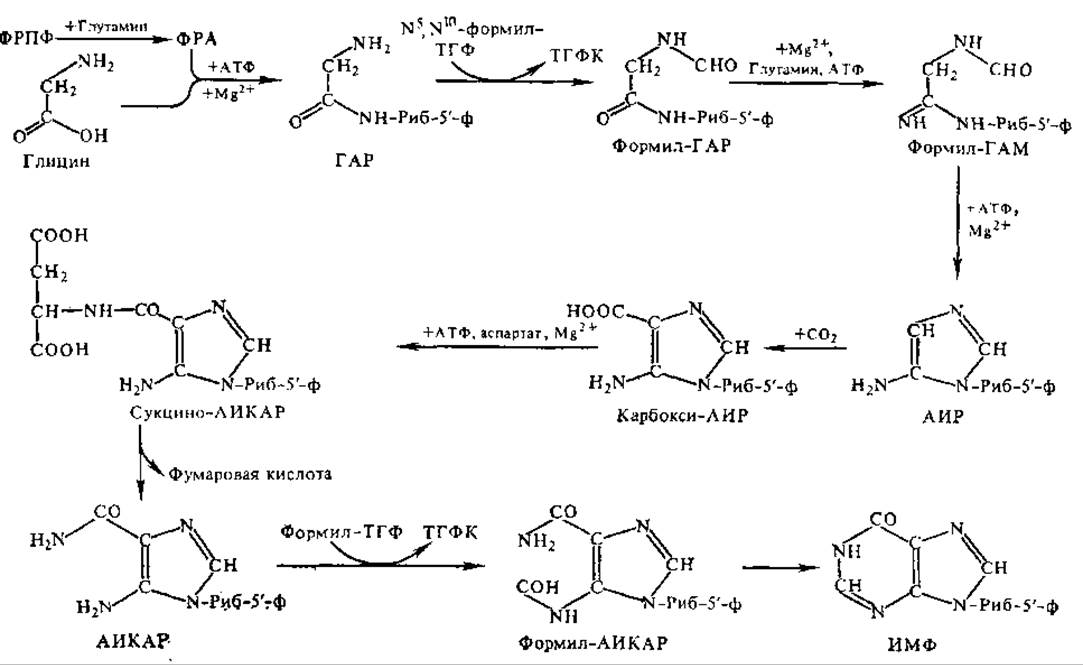
Рис. 4.11. Схема биосинтеза пуриновых нуклеотидов:
АИКАР — 5-аминоимидазол-4-карбоксамидрибонуклеотид, АИР — 5- аминонмидазолрибонуклеотид, ГАР — глицинамидрибонуклеотид, ИМФ — инозин-5' монофосфат, Риб-5-ф — рибозо-5-фосфат, N6,N10-ТГФ — N-форм илтетрагидрофолиевая кислота. ТГФК — тетрагидрофолиевая кислота, формил-ГAM — формилглицинамидинрибонуклеотид, ФРА — 5- фоcфорибозилaмин, ФРПФ — 5-фосфорибозил-1-пирофосфат
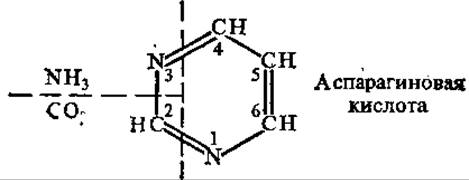
Рис. 4.12. Происхождение атомов пиримидинового кольца
4.5.3. Биосинтез пиримидиновых нуклеотидов. Применение метода изотопов позволило выяснить происхождение отдельных звеньев пиримидинового ядра (рис. 4.12).
Первым этапом в синтезе пиримидиновых нуклеотидов является образование карбамоилфосфата из NH3 и СО2. Карбамоилфосфат вступает в реакцию с асп и под действием аспартат-карбамоилтрансферазы превращается в карбамоиласпартат. Последний подвергается циклизации и окислению, в результате чего образуется оротовая кислота (см. разд. 10.4.1), т. е. завершается формирование пиримидинового кольца.
Таким образом, в процессе синтеза пиримидиновых нуклеотидов в отличие от рассмотренного выше синтеза пуриновых нуклеотидов сначала образуется свободное азотистое основание. Только после этого протекает пирофосфорилазная реакция, в ходе которой к оротовой кислоте присоединяется рибозо-5'-фосфат от фосфорибозилпирофосфата (ФРПФ), образуется нуклеотид оротидин-5'-монофосфат (ОМФ). Декарбоксилирование ОМФ приводит к образованию уридин-5'-монофосфата (УМФ) — предшественника других пиримидиновых нуклеотидов (рис. 4.13).
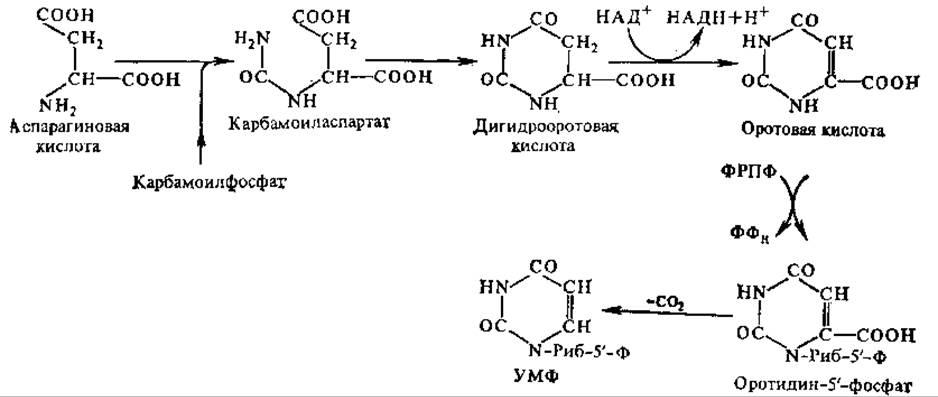
Pис. 4.13. Схема биосинтеза пиримидиновых нуклеотидов (пояснение см. в тексте)
Превращение урацила в цитозин происходит на уровне нуклеозидтрифосфатов. Оно осуществляется под действием фермента ЦТФ-синтетазы: УТФ + NH3 + АТФ→ЦТФ + АДФ + Фн. Тиминовые нуклеотиды образуются в результате метилирования дезоксиуридинмонофосфата. Реакция катализируется ферментной системой, которую часто называют тимидилат-синтазой. Процесс весьма сложен и протекает в несколько стадий. Источником одноуглеродного фрагмента при С-5 служит кофермент N5,N10-метилентетрагидрофолиевая кислота.
4.5.4. Биосинтез дезоксирибонуклеотидов. Превращение рибозы в дезоксирибозу происходит на нуклеотидном уровне. Механизм этого процесса был выяснен при изучении восстановления рибонуклеотидов в экстрактах Е. coli. Фермент рибонуклеозид-дифосфат-редуктаза катализирует восстановление всех четырех рибонуклеозиддифосфатов — АДФ, ГДФ, ЦДФ, УДФ в их дезоксипроизводные дАДФ, дГДФ, дЦДФ, дУДФ. Источник восстанавливающей способности фермента in vivo неизвестен.
In vitro активны два донора водорода. Один из них — низкомолекулярный серосодержащий белок тиоредоскин, две SH-группы которого окисляются с образованием дисульфидного мостика. Вторым донором водорода может служить восстановленный глутатион. Редуктазные системы, выделенные из различных животных клеток, сходны с описанной системой Е. coli.