Основы биохимии - А. А. Анисимов 1986
Нуклеиновые кислоты
Синтез РНК
4.8.1. Транскрипция. Транскрипция у прокариот. Транскрипция — процесс, посредством которого заключенная в ДНК генетическая информация «переписывается» в одиночные цепи РНК. Впоследствии РНК переносится к рибосомам.
Единицы процесса транскрипции несут информацию о структуре одного или нескольких белков. Участок ДНК, в котором заключена информация о структуре одного белка, называется цистроном или структурным геном. Регуляция транскрипции осуществляется благодаря наличию в ДНК специальных регуляторных участков. Регуляторная зона включает в себя промотор, оператор, а нередко и другие участки управления (рис. 4.17).
Промотор содержит участок первоначального прочного связывания ДНК-зависимой РНК-полимеразы с ДНК. Оператор — регуляторный участок, который связывается с репрессорами — белками, контролирующими синтез мРНК в соответствии с потребностями клетки. Оператор и промотор иногда частично перекрываются. В некоторых единицах транскрипции (опероны) между оператором и структурными генами располагается так называемая лидерная зона. Она транскрибируется, но, как правило, не транслируется. В ее границах располагаются участок связывания рибосомы на мРНК и аттенюатор. Последний регулирует транскрипцию, оказывая влияние на связь РНК-полимеразы с матрицей ДНК. После структурных генов находится терминатор.
Последовательность нуклеотидов ДНК, ограниченная промотором и терминатором, кодирующая одну молекулу мРНК и контролируемая оператором, называется опероном. У прокариот известны опероны, в состав которых входит несколько цистронов (генов), кодирующих структуру ферментов одной метаболической цепи. Благодаря наличию регуляторной зоны все пистроны включаются и выключаются одновременно. Представления об опероне были сформулированы в 1959 г. Ф. Жакобом и Ж. Моно и являются основой понимания механизмов управления работой генов у прокариот. В геноме находятся также единицы транскрипции, не контролируемые каким-либо оператором, функционирующие конститутивно.
ДНК-зависимый синтез РНК можно разбить на несколько стадий, которые в целом составляют цикл транскрипции. Они подробно изучены на прокариотических объектах. Первая стадия транскрипции — инициация — включает взаимодействие РНК-полимеразы с матричной ДНК. РНК-полимераза может связываться с любым участком ДНК, при этом образуется неспецифический слабый комплекс с коротким периодом полураспада. Через серию актов ассоциации — диссоциации на случайных последовательностях ДНК РНК-полимераза находит промотор. В области промотора образуется сначала закрытый стабильный комплекс ДНК и РНК-полимеразы. Затем происходит локальная денатурация ДНК. РНК- полимераза получает прямой доступ к основаниям ДНК, образуется открытый комплекс.
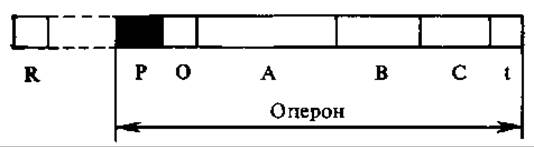
Рис. 4.17. Строение оперона прокариот:
Р — промотор, О — оператор, А, В, С — цистроны, t — терминатор, R — ген-регулятор
Взаимно комплементарные цепи молекулы ДНК антипараллельны, комплементарная вырожденность кода отсутствует, поэтому обе цепи ДНК не могут кодировать один белок. Считают, что транскрибируется или только одна из двух цепей, или обе, но лишь одна из образовавшихся молекул РНК является матричной, вторая же выполняет иные функции. Цепь ДНК, комплементарную мРНК, обычно называют кодирующей, вторую цепь — замыкающей. Так как транскрипция несимметрична (идет на одной цепи из двух), можно предполагать, что и промотор фиксирован лишь в одной из двух цепей ДНК, имеет тоже несимметричную структуру и определяет направление движения РНК-полимеразы, т. е. выбор ею кодирующей цепи.
Если имеются соответствующие рибонуклеозидтрифосфаты, то начинается синтез РНК. Первыми нуклеотидами при инициации транскрипции почти всегда бывают А или Г. Образуется трехкомпонентный комплекс ДНК-РНК-полимеразарастущая цепь РНК. Промотор не транскрибируется. Оператор в некоторых случаях частично транскрибируется.
Элонгация (рост цепи) РНК происходит в направлении Рибонуклеотиды присоединяются к 3'-ОН-концу последовательно, один за другим, соответственно матрице ДНК. Скорость процесса элонгации в клетках Е. coli in vivo составляет 45—50 нуклеотидов при 37°С в 1 с.
Терминацию синтеза РНК вызывает последовательность нуклеотидов в ДНК — терминатор или стоп-сигнал. Структура терминаторов полностью не выяснена. Известно, что в их состав входят длинные блоки. У Е. coli обнаружены стоп-сигналы другого типа, которые действуют только в присутствии белка, называемого p-фактором. Этот белок с М = 50 000, обычно не связанный с РНК-полимеразой.
При его участии цепь РНК освобождается от матрицы ДНК, но фермент еще остается связанным с ДНК. Для высвобождения фермента нужны некоторые дополнительные факторы. Выделен еще один белок, участвующий в терминации, — каппа-частица.
В клетках бактерий к готовому, начинающему отделяться от матрицы участку мРНК присоединяются рибосомы. Они способствуют отделению мРНК от матрицы и начинают синтез белка. Так образуется единый транскрипционно - трансляционный комплекс, который удалось обнаружить методом электронной микроскопии (рис. 4.18).

Рис. 4.18. Транскрипция и трансляция в клетках Е. coli (согласно данным электронной микроскопии).
На двухцепочечной ДНК растут цепи мРНК. Сверху, где транскрипция только началась, цепи мРНК короче, а ниже, где транскрипция продолжалась в течение определенного времени, — длиннее. К цепям мРНК присоединяются рибосомы, начинается синтез полипептидных цепей
Особенности транскрипции у эукариот. Если у прокариот транскрипция и трансляция сопряжены, то у эукариот эти процессы разобщены во времени и пространстве. Важная роль в этом принадлежит ядру клетки, разграничивающему аппараты транскрипции и трансляции. При описании транскрипции у эукариот термин «оперон» может быть использован весьма условно, так как гены, детерминирующие структуру белков одной метаболической цепи, не обязательно расположены рядом, а могут быть локализованы даже в разных хромосомах. В связи с этим мутации, прерывающие один и тот же метаболический путь, у эукариот также бывают «разбросаны» по всему геному. Для структурных генов эукариот выявлены не одиночные регуляторные участки, а их большие серии. Также отличаются ферментные системы, считывающие информацию с генома про- и эукариот, и этапы превращений первичных продуктов транскрипции в зрелые молекулы мРНК.
Большинство цитоплазматических молекул РНК синтезируется первоначально в ядре в виде высокомолекулярных предшественников — пре-мРНК и пре-рРНК. Они гораздо длиннее, чем цитоплазматические РНК, подвергаются процессингу (созревание). Транскрипция у эукариотических организмов осуществляется примерно по тем же стадиям, что и у прокариот, но ее проводят три разных РНК-полимеразы. Транскрипция РНК-полимеразами І и III мало изучена. Однако установлено, что инициация транскрипции в разных генах определяется их внутренними участками, имеющими сходное строение, но расположенными на различном расстоянии от места начала транскрипции.
РНК-полимераза II эукариотических клеток катализирует синтез всех пре-мРНК. Механизм функционирования РНК-полимеразы II в значительной степени напоминает прокариотические системы. Изучение продуктов транскрипции позволило выявить характерные особенности строения как структурных, так и регуляторных зон генома. Наиболее детально изучены гистоновые гены и в несколько меньшей мере — глобиновые, овальбуминовый и кональбуминовый.
У большинства генов этой группы обнаружена сходная последовательность (ТАТААА или более короткая — ТАТА или АТА), названная последовательностью Гольдберга-Хогнесса. Она расположена «вверх по течению» от точки начала транскрипции на 21— 28 нуклеотидных пар. С обеих сторон от АТ-богатой последовательности располагаются ГЦ-богатые участки. Предполагают, что ТАТА-участок важен для инициации транскрипции, т. е. выполняет роль промотора. ТАТА-блок определяет выбор точки начала транскрипции, но не влияет на ее эффективность, поэтому иногда его называют селектором. На определенном расстоянии от ТАТА-последовательности располагается инициаторная область, с которой непосредственно начинается транскрипция мРНК, она заканчивается до кодона АТГ. На расстоянии примерно 100—300 пар нуклеотидов от точки начала транскрипции за селектором располагается другой функционально важный участок — модулятор (модуляторов может быть несколько). Он определяет скорость синтеза неизменных по структуре молекул мРНК только в присутствии селектора и инициатора (рис. 4.19).
На инициацию транскрипции могут влиять и более отдаленные области генома. По-видимому, истинный промотор генов, транскрибируемых РНК-полимеразой ІІ, формируется из нескольких участков генома за счет определенной конформации хроматина. За точность и эффективность транскрипции отвечают также многочисленные белковые факторы клетки, которые еще плохо изучены.
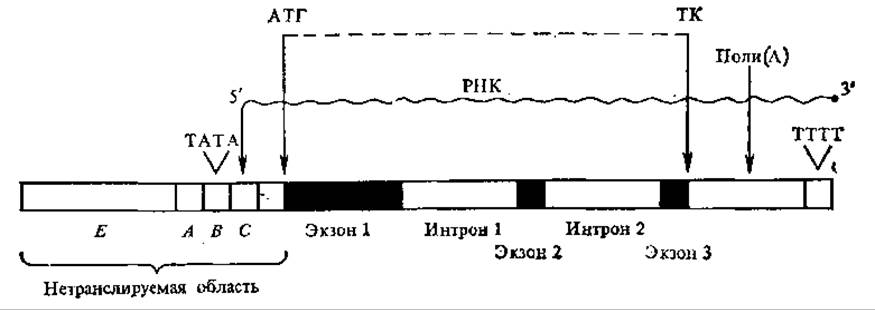
Рис. 4.19. Строение единицы транскрипции эукариот:
А и В — модуляторы, В — селектор. С — инициатор, t — терминатор транскрипции, поли (А)-участок, являющийся, видимо, затравкой при полиаденилировании в процессинге; 5' — первый транскрибируемый кодон; АП — кодон, инициирующий трансляцию на рибосоме; ТК — кодон, терминирующий трансляцию; темными участками обозначены экзоны, светлыми — нитроны
Структурные зоны генов эукариот также имеют некоторые особенности, отличающие их от структурных генов бактерий. Внутри участков, кодирующих структуру белков, существуют разрывы — интроны. Они чередуются с экзонами — участками гена, которые кодируют последовательности зрелых мРНК (рис. 4.19). Первый экзон, если он не кодирует полипептид, а является матрицей для 5'-нетранслируемого участка мРНК, называется лидерной последовательностью. Число экзонов сильно варьирует у разных генов. Существует предположение, что экзоны в ряде случаев соответствуют определенным доменам в кодируемых белках. Например, это показано в исследованиях иммуноглобулиновых генов. Возможно, перестановка крупных блоков (экзонов) в генах является одним из путей эволюции последних.
Терминация транскрипции в эукариотических клетках изучена недостаточно. Существуют, например, сведения, полученные при анализе гистоновых генов. В области, примыкающей к 3'-концу структурной зоны гена, на расстоянии 23—47 пар нуклеотидов от ее последнего кодона, обнаружен консервативный участок, состоящий из двух коротких палиндромов, которые разделены последовательностью ТТТТ. Предполагают, что РНК образует в этом участке «шпильку», которая участвует в терминации.
Работы американского цитолога О. Миллера (1969) позволили увидеть единицы процесса транскрипции на электронных микрофотографиях. Из яйцеклеток амфибий ученым были выделены действующие гены, кодирующие рибосомную РНК. На схеме, составленной по микрофотографии, изображены нити ДНК с прикрепленными к ним молекулами РНК-полимеразы и свисающими с молекул полимеразы нитями РНК (рис. 4.20). Схема показывает, что транскрипцию проводят сразу 80—100 молекул РНК-полимеразы, которые последовательно связываются с промотором.

Рис. 4.20. Образование молекул РНК на матрице ДНК, соответствующей одному гену
4.8.2. Процессинг мРНК. В эукариотических клетках первичные транскрипты превращаются в мРНК в ходе процессинга. Пре-мРНК составляют основную фракцию гетерогенной ядерной РНК (гяРНК), которую выделили впервые в 1961 г. Г. П. Георгиев и сотрудники. Пре-мРНК содержит от 5 000 до 50 000 нуклеотидов, в то время как мРНК относительно коротки, их средний размер составляет около 2000 нуклеотидов. Одна мРНК кодирует одну полипептидную цепь, т. е. является моноцистронной. Каждая молекула пре-мРНК обычно дает начало только одной молекуле мРНК, при этом большая часть цепи пре-мРНК (иногда до 90% и более), соответствующая некодирующей зоне ДНК, подвергается ферментативному расщеплению до свободных нуклеотидов и в цитоплазму не поступает. Эти изменения представляют собой часть процессинга.
Процессинг (посттранскрипционная модификация) включает в себя: 1) отрезание «лишних» концевых последовательностей, 2) расщепление длинных первичных транскриптов, «вырезание» из них участков, транскрибированных с интронов, 3) добавление нуклеотидов к 3'-концу транскрипта, 4) добавление нуклеотидов к 5'-концу транскрипта, 5) модификацию оснований в транскрипте.
Процессинг протекает в ядре. К 3'-концу пре-мРНК присоединяется (с участием специального фермента) последовательность из 150—200 адениловых нуклеотидов — поли (А)-фрагмент, к 5'-концу присоединяется кэп (см. разд. 4.4.3). Вырезание интронов из первичного транскрипта сопровождается сплайсингомМеханизм сплайсинга точно не выяснен. Предполагают, что в нем участвуют особая UI-РНК и некоторый фермент. [UI-РНК узнает места соединения интронов и экзонов, она взаимодействует с ними по принципу комплементарности, образуя гибрид пре-мРНК — комплекс Ul- РНК-фермент. Это приводит к сближению двух участков пре-мРНК, транскрибированных с соседних экзонов.
Участок, транскрибированный с нитрона, образует петлю (рис. 4.21). Образовавшиеся гибриды могут быть местами атак РНКаз, специфичных для двухнитевых участков РНК. После вырезания «лишнего» участка, т. е. транскрибированного с интрона, специальные лигазы сшивают два конца разрезанной молекулы пре-мРНК.
В результате процессинга из пре-мРНК получается молекула мРНК, имеющая 5'-кэп, универсальный Заполи (А) блок и одну кодирующую последовательность. Для прокариотических клеток процессинг не характерен.
Сплайсинг протекает в эукариотических клетках при биосинтезе как мРНК, так и тРНК, рРНК. Например, сплайсингу подвергаются некоторые тРНК дрожжей, хотя их интроны очень малы по размерам. У дрозофилы гены рРНК также содержат интрон.
4.8.3. Рибонуклеопротеиновые комплексы (PHП-комплекеы). Молекулы РНК сразу же после транскрипции соединяются с белками, образуя компактные структуры. РНП-комплексы, содержащие гяРНК, называются ядерными информосомами (гяРНП). Отношение белок : РНК составляет в них 4:1.
Кроме гяРНК в составе гяРНП обнаружены низкомолекулярные стабильные РНК, имеющие коэффициенты седиментации от 4,5 S до 6,5 S. Они тесно связаны как с самими молекулами гяРНК, так и с белками РНП-частиц.
Общую схему строения гяРНП можно представить следующим образом. Вдоль цепи гяРНК формируются многочисленные моночастицы, расположенные нестрого регулярно. Эти 30—40S частицы состоят из белковой глобулы (информофера), на поверхности которой некоторым образом «накручен» небольшой (600 нуклеотидов) участок цепи гяРНК. Моночастицы чередуются с гетерогенными (30—200S) РНП-комплексами. Такие структуры обнаруживаются при электронной микроскопии в виде отходящих от хроматина фибрилл со множеством повторяющихся РНП-частиц.
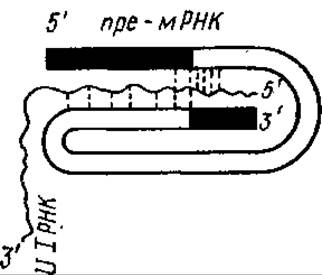
Рис. 4.21. Предполагаемая структура, образующаяся при сплайсинге.
Пунктиром обозначены водородные связи, образовавшиеся между комплементарными азотистыми основаниями. Темными участками представлены последовательности пре-мРНК, транскрибированные с экзонов, светлыми — с интронов
1 Слово сплайсинг (splicing) взято из английской морской терминологии, где так называют сращивание канатов без узла.
Предполагают, что определенные третичная и четвертичная структуры гяРНК, обусловленные взаимодействием с белками, способствуют точному протеканию процессинга и сплайсинга. Установлено, что процессинг гяРНК протекает в составе РНП-частиц. В цитоплазме мРНП появляются после процессинга гяРНП. Здесь обнаруживаются свободные мРНП-частицы (цитоплазматические информосомы), или мРНП, связанные с полисомами (полисомные информосомы). В них отношение белка и РНК по массе составляет 3: 1—4: 1. В состав мРНП-частиц входят молекулы мРНК разных размеров. Связанные с полисомами мРНК активно транслируются. В полисомных и свободных цитоплазматических информосомах содержится не более 15—20 различных белков.
В полисомных информосомах обнаруживаются два основных белка. В свободных информосомах содержится больше белков, среди них — ингибирующие трансляцию белки. Белки мРНП существенно отличаются от белков гяРНП. Переход из ядра в цитоплазму сопровождается сменой белкового компонента РНП-частиц. Белки, связывающиеся с информосомами, маскируют мРНК и обеспечивают ее хранение в цитоплазме в нетранслируемом состоянии. Переход мРНП из информосом в полисомы также должен сопровождаться изменением в составе белков: отщеплением или модификацией репрессорных и связыванием активаторных белков (факторы инициации и др.).
А. С. Спирин (1966) высказал предположение, что информосомы представляют собой форму хранения мРНК в цитоплазме до вхождения мРНК в полисомы и трансляции. В таком виде, например, хранится в неактивном состоянии мРНК яйцеклетки до оплодотворения, в семенах до прорастания.
Таким образом, в эукариотических клетках мРНК всегда находится в комплексе с белками. РНП-комплекс — это единственная форма существования мРНК в животной и растительной клетке, от момента синтеза пре-мРНК в ядре до распада мРНК в цитоплазме.
4.8.4. Синтез рибосомных и транспортных РНК. Все рибосомные и транспортные РНК синтезируются на матрице ДНК. У Е. coli рибосомные РНК образуются в виде большого предшественника 30 S. Он дает начало всем рРНК — 16 S, 23 S, 5 S. При созревании рРНК интенсивно метилируются. Транспортные РНК у прокариот также транскрибируются в виде предшественников, содержащих одну или более тРНК. Например, гены фага Т4 кодируют предшественник сразу двух тРНК — пролиновой и сериновой. Он «нарезается» на мономеры, от которых отщепляются «лишние» участки. Если у продуктов расщепления отсутствует ЦЦА-последовательность, то она достраивается специальным ферментом.
В эукариотических клетках рРНК кодируются областью хромосомы, которая образует ядрышко — ядрышковый организатор. Здесь несколько сотен раз повторяется участок, который кодирует 28 S, 18 S, 5,8 S pPHK. На транскрибируемой области РНК-полимераза I образует крупный предшественник. От него чрезвычайно быстро удаляется несколько сотен нуклеотидов, находящихся на 5'-конце. Оставшийся предшественник подвергается процессингу. В клетках млекопитающих обнаружено 8 различных по размеру пре-pPHK (от 45 S до 12 S), поэтому предполагают, что существуют по крайней мере три пути процессинга. Места разрывов предшественника (45 S РНК) одинаковы во всех случаях, мест разрывов всего пять, но порядок расщепления может отличаться. Обычно 45 S расщепляется на две части эндонуклеазой. Затем другие ферменты удаляют спейсеры (разделяющие последовательности). Из одной половины образуется 18 S рРНК, из другой — 28 S и 5,8 S рРНК, связанные водородными связями.
Синтез 5 S рРНК и тРНК катализирует РНК-полимераза III. Пре-5 S рРНК содержит небольшое число «лишних» нуклеотидов на 3'-конце молекулы, они удаляются при процессинге. В предшественниках тРНК имеются избыточные последовательности, которые могут быть расположены как на обоих концах молекулы, так и внутри ее. При процессинге пре-тРНК эти избыточные последовательности отщепляются, идет модификация оснований, «вырезается» (если существует) вставочная последовательность, происходит сплайсинг. К 3'-концу пре-тРНК присоединяется последовательность ЦЦА. Все эти процессы осуществляются в ядре.