Основы биохимии - А. А. Анисимов 1986
Интеграция и регуляция метаболизма
Гормоны человека и животных
12.6.1. Общая характеристика, механизмы действия. К гормонам (от греч. гормао — привожу в движение, побуждаю) относятся органические соединения, вырабатываемые железами внутренней секреции (эндокринные железы), транспортируемые кровью к клеткам-мишеням и активно влияющие на метаболические, морфогенетические и физиологические процессы. Однако некоторые гормоны синтезируются не только эндокринными железами, но и в клетках других органов и тканей. Так, гормоны поджелудочной железы (инсулин и глюкагон) образуются также и в клетках кишечника, катехоламины (гормоны надпочечников) синтезируются и в нервных окончаниях.
Существенной отличительной особенностью гормонов от других биологически активных веществ является специализация клеток, их синтезирующих. Железы внутренней секреции не содержат выводных протоков, их клетки оплетены обильной сетью кровеносных и лимфатических капилляров, выделение продуктов жизнедеятельности происходит непосредственно в просвет этих сосудов. Этим эндокринные железы отличаются от экзокринных, которые выделяют свои секреты через выводные протоки.
Действие гормонов на органы и ткани характеризуется рядом особенностей.
1. Высокой биологической активностью, проявляющейся в том, что гормоны оказывают физиологическое действие в очень малых концентрациях.
2. Специфичностью действия: гормоны вызывают строго специфические ответные реакции органов и тканей.
3. Дистантностью действия: гормоны, как правило, переносятся кровью далеко от места их образования и оказывают действие на определенные чувствительные к ним органы-мишени или клетки-мишени (только в последнее время обнаружены исключения из этого правила — некоторые гормоны могут действовать в месте своего образования).
4. Относительно небольшой период полужизни, обычно меньше 1 ч; в результате этого эффективное функционирование гормонов возможно при непрерывном синтезе и секретировании их в течение всего требуемого времени при данном состоянии организма.
Эндокринные железы появились у членистоногих и позвоночных как один из основных компонентов нейроэндокринной регуляторной системы для поддержания гомеостаза. Гормоны или вещества, близкие к ним по действию, в последнее десятилетие были обнаружены у червей.
Гормоны млекопитающих по химическому строению можно разделить на три большие группы: 1) белки и пептиды, 2) производные аминокислот, 3) стероиды.
В особую группу обычно выделяют так называемые тканевые гормоны или гуморальные факторы, которые образуются не в эндокринных железах, а во многих тканях организма (гистамин, простагландини и др.).
Ряд полипептидных гормонов синтезируется эндокринными железами в виде более крупных молекул предшественников — прогормонов, в некоторых случаях продукты рибосомального синтеза представляют собой еще более крупные белки — препрогормоны. Последние кроме пептида-ингибитора (как все прогоморны) содержат дополнительный пептид, определяющий транспорт гормона через мембрану при его секреции железой.
Существуют представления о двух основных механизмах действия гормонов (стероидных и нестероидных). Специфичность гормонов по отношению к клеткам-мишеням связана с наличием белковых рецепторов. Рецепторы большинства гормонов (полипептидного строения и производных аминокислот, т. е. нестероидные) находятся в плазматической мембране клеток. Ответная реакция возникает при связывании гормона рецептором. Так как рецепторы являются гликопротеинами, их специфичность обусловлена углеводным компонентом. Определенную роль в этом играют и углеводные фрагменты ганглиозидов, находящихся в липидном бислое плазматической мембраны.
Взаимодействие нестероидного гормона с рецептором активирует аденилатциклазу, что приводит к увеличению концентрации внутриклеточной цАМФ, а следовательно, фосфорилированию протеинкиназами различных белков: ферментативных, мембранных, рибосомальных, хроматиновых. В результате могут возникнуть самые разнообразные изменения в метаболизме клетки. Фосфорилирование белков хроматина оказывает влияние на транскрибирование генов, контролируя тем самым скорость синтеза ферментов и других белков. Исключение в группе гормонов-полипептидов и производных аминокислот составляют инсулин и тиреоидные гормоны. Рецепторами последних являются внутриядерные белки, входящие в состав хроматина. Рецепция приводит к усилению синтеза РНК в клетках, активации РНК-полимеразы, увеличению скорости синтеза белков. Видимо, тиреоидные гормоны стимулируют последний на стадии транскрипции, что и лежит в основе их действия на рост, развитие и дифференцировку клеток. Кроме того, тиреоидные гормоны содержат рецепторы в митохондриальной мембране, в результате чего оказывается влияние на энергетический обмен.
Рецепторы инсулина локализованы на поверхности клеток-мишеней, гормон проявляет свое действие, не проникая в клетку. Однако большинство эффектов инсулина (влияние на липолиз, синтез гликогена и др.) противоположно действию цАМФ. Следовательно, сходства инсулина в этом отношении с другими гормонами-полипептидами нет: он не стимулирует аденилатциклазную активность. Кроме того, специфические инсулинсвязывающие рецепторы обнаружены в ядрах некоторых клеток, но роль их недостаточно выяснена. Связывание инсулина с клетками-мишенями приводит к увеличению скорости синтеза и накопления белков, а также гликогена и липидов (т. е. к запасанию энергии).
Стероидные гормоны проникают в цитоплазму клеток-мишеней и связываются с определенными цитоплазматическими белками-рецепторами. Образовавшиеся комплексы перемещаются в ядра клеток и присоединяются к хроматину. Изменяя доступность для транскрипции определенных участков ДНК, стероидные гормоны влияют на синтез мРНК, т. е. действуют на уровне генома.
Таким образом, механизм действия гормонов направлен в основном на белки: на скорость их синтеза и активацию, модификацию уже синтезированных белков. Стероидные и тиреоидные гормоны осуществляют регуляторную функцию первым путем, влияя на синтез белка в стадии транскрипции. Гормоны-белки, пептиды и производные аминокислот (кроме тиреоидных) воздействуют на метаболизм прежде всего через активацию и модификацию белков в результате их протеинкиназного фосфорилирования (второй путь). В некоторых случаях вследствие фосфорилирования хроматиновых и рибосомальных белков нестероидные гормоны тоже могут (по уже опосредованно) влиять и на сам синтез белков.
Между железами внутренней секреции складываются сложные взаимодействия. Это связано с тем, что: 1) на функцию каждого органа оказывает влияние сразу несколько гормонов; 2) гормоны, вырабатываемые одними железами, оказывают влияние на функцию других эндокринных желез. Выделяют три основных типа взаимодействия между эндокринными железами.
1. Взаимодействие по принципу положительной прямой или отрицательной обратной связи. Так, передняя доля гипофиза вырабатывает тиреотропный гормон, стимулирующий образование гормонов щитовидной железы, — положительная прямая связь. С другой стороны, повышение уровня гормонов щитовидной железы выше нормы тормозит образование тиреотропного гормона гипофиза — отрицательная обратная связь.
2. Синергизм гормональных влияний. Например, как адреналин — гормон мозгового слоя надпочечников, так и глюкогон а-клеток островкового аппарата поджелудочной железы активируют распад гликогена печени до глюкозы и вызывают увеличение содержания глюкозы в крови.
3. Антагонизм гормональных влияний. Примером могут служить женские половые гормоны — эстрогены и прогестерон. Первые усиливают сократительную функцию матки, второй — тормозит ее.
Существует хорошо выраженная связь между эндокринной и нервной системой. В секреторных клетках гипоталамуса (отдел головного мозга) образуются гормоны — низкомолекулярные полипептиды, названные рилизинг-факторами или регуляторными факторами. Их органом-мишенью является аденогипофиз (передняя доля гипофиза), Рилизинг-факторы, вырабатываемые в гипоталамусе, по мнению одного из ведущих советских эндокринологов Н. А. Юдаева (1976), можно рассматривать как универсальные химические сигналы, посредством которых осуществляется передача нервных импульсов на эндокринную систему.
Нервные стимулы, поступающие из таламуса и коры мозга, влияют на скорость высвобождения гипоталамических регуляторных гормонов, которые попадают в аденогипофиз по гипоталамо- гипофизарной портальной системе циркуляции крови и вызывают секрецию определенных гормонов аденогипофиза. Так, с участием указанных нервных путей осуществляется быстрая регуляция аденогипофизарной секреции в ответ на различные воздействия внешней среды (холод, гипоксия, травмы, яды) или на определенное физиологическое состояние (страх, тревога).
Комиссия по биохимической номенклатуре ШРАС Международного биохимического общества предложила названия тех гипоталамических рилизинг-факторов, которые способствуют образованию гормонов гипофиза, оканчивать словом либерин, а ингибирующих — статин (например, кортиколиберин, пролактостатин).
Гипоталамус непосредственно связан (анатомически, функционально) и с нейрогипофизом (задняя доля гипофиза). Секретируемые нейрогипофизом гормоны (в ответ на определенные нервные стимулы) синтезируются в гипоталамусе, мигрируют в составе гранул по нервным волокнам и накапливаются у их окончаний в нейрогипофизе. Все изложенное свидетельствует о том, что «гипоталамо-гипофизарная ось» является анатомической и биохимической основой интеграции действия нервной системы и эндокринной.
В свою очередь гормоны оказывают влияние на нервную систему. При этом происходит изменение метаболизма нейронов мозговой ткани — обмена электролитов и аминокислот, процессов образования и связывания аммиака. Влияние гормонов на функции нейронов мозговых структур приводит к их участию в регулирующем и коррегирующем воздействии ЦНС на различные органы. Гормоны оказывают действие и на уровне рецепторного аппарата, особенно на хеморецепторы, расположенные в стенках кровеносных сосудов.
12.6.2. Щитовидная железа. Секретирует тироксин и трииодтиронин, а также кальцитонин (в особых парафолликулярных клетках). Исходными продуктами синтеза первых двух гормонов являются тирозин и неорганический иод.
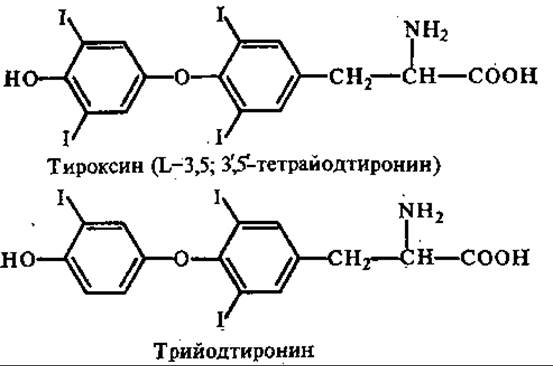
Синтез тироксина и трииодтиронина катализируется тиреоидпероксидазой путем иодирования остатков тирозина в составе молекулы белка тиреоглобулина. Последний не обладает гормональными свойствами, они присущи только иодтиронинам, освобождающимся при его протеолизе, который осуществляется внутриклеточными протеиназами эпителиальных клеток. Синтез и протеолиз тиреоглобулина происходит непрерывно.
Тироксин связывается в плазме крови с белковыми носителями: тироксинсвязывающим глобулином, преальбумином и альбумином.
Гипофункция щитовидной железы (гипотиреоз) сопровождается снижением основного обмена, температуры тела, слизистыми отеками. Это заболевание называется микседемой. Если гипотиреозом страдают дети, развивается кретинизм, сопровождающийся задержкой роста и умственного развития. Гиперфункция щитовидной железы характеризуется резким усилением обмена, учащением пульса, дрожанием рук, пучеглазием. Этот комплекс получил название базедовой болезни.
Кальцитонин представляет собой линейный полипептид, построенный из 32 остатков аминокислот. Все 32 остатка существенны для проявления биологической активности. Первичная структура кальцитонинов различных видов животных неидентична, но довольно близка. Так, кальцитонин трески отличается от такового высших позвоночных только по девяти аминокислотным остаткам. Основная физиологическая роль кальцитонина состоит в том, что он предупреждает возможное повышение концентрации Са2+ в крови путем торможения процесса выхода Са2+ из костей.
12.6.3. Паращитовидные железы. Секретируют паратгормон — белок (М9500), находящийся в паращитовидных железах в виде прогормона, на N-конце которого содержится дополнительный гексапептид. Предполагают, что отщепление последнего приводит к появлению биологической активности.
Главная функция паратгормона — регуляция концентрации ионов Са в крови (путем ее увеличения после падения по какой-либо причине). Основное место действия паратгормона — почки и кости скелета.
12.6.4. Половые железы. Семенники секретируют тестостерон и дигидротестостерон (андрогены) в количестве 5—7 мг/сут.

Биосинтез этих гормонов начинается с холестерина. Андрогены обусловливают нормальный рост мужских половых органов, развитие вторичных половых признаков у мужчин. При недостатке тестостерона замедляется биосинтез белков, развивается ожирение, утрачивается волосяной покров. Андрогены оказывают выраженное анаболическое действие на обмен азота и кальция.
Женские половые гормоны (эстрогены) синтезируются главным образом в яичниках. Фолликулы яичников секретируют наиболее активный эстроген — ß-эстрадиол в количестве 1 мг/сут.

Он обеспечивает развитие половых органов и вторичных половых признаков. При недостаточности эстрадиола нарушаются циклы менструации, происходят самопроизвольные выкидыши, развивается ожирение. Эстрогены оказывают выраженное влияние на синтез белков, нуклеиновых кислот, обмен липидов.
12.6.5. Желтое тело. Продуцирует прогестерон и релаксин. Прогестерон образуется во второй половине менструального цикла. Особенно много его образуется во время беременности. Так как он воздействует на систему гиалуронидаза — гиалуроновая кислота, прогестерон необходим при подготовке слизистой оболочки матки для закрепления в ней оплодотворенного яйца. Прогестерон влияет на проницаемость клеточных и митохондриальных мембран на процесс окислительного фосфорилирования, образование мРНК.

Релаксин — полипептид (М6000), состоящий из двух неидентичных цепей, соединенных дисульфидными мостиками (большое сходство в строении молекулы с инсулином). Релаксин подготавливает матку и тазовые сочленения к родам.
12.6.6. Надпочечники. Состоят из мозгового вещества и коркового слоя. Гормонами мозгового вещества являются адреналин и норадреналин. Эти гормоны и близкие к ним амины, содержащие в своей структуре катехол, называются катехоламинами. Роль адреналина и продуктов его превращения в организме были глубоко исследованы в работах А. М. Утевского (1950).

Основной путь синтеза катехоламинов: тирозин→диоксифенилаланин (ДОФА)→диоксифенилэтиламин (ДОФАМИН)→норадреналин→адреналин.
Адреналин и норадреналин, стимулируя аденилатциклазу в органах-мишенях, активируют фосфоролиз гликогена в печени и скелетных мышцах, резко повышая уровень глюкозы в крови; увеличивают силу сердечных сокращений, секрецию амилазы слюнных желез. Катехоламины обладают выраженной липидмобилизующей активностью.
Физиологическое действие адреналина и норадреналина сходно с влиянием симпатических нервов (например, повышение кровяного давления), возбуждение этих нервов приводит к секреции указанных гормонов, поэтому последние называют симпатомиметическими аминами. В связи с тем, что они являются не только гормонами, но и нейромедиаторами, эти амины могут синтезироваться в нервной системе из тирозина по тому же пути, что и в надпочечниках. В крови животных и человека в спокойном состоянии содержится очень мало адреналина. Резкое эмоциональное возбуждение, боль, физическое напряжение, охлаждение приводят к значительному повышению его концентрации в крови, так как он служит инициатором «мобилизационной готовности организма».
В целом вся симпато-адреналовая система (мозговой слой надпочечников и симпатический отдел нервной системы) обеспечивает готовность организма к защитным реакциям, требующим активной двигательной деятельности. Эта готовность выражается в усилении функции сердечно-сосудистой системы, повышении энергетического баланса, торможении деятельности желудочно-кишечного тракта, усилении кровоснабжения скелетной мускулатуры. В связи с этим адреналин называют «гормоном борьбы и бегства». У животных значение симпато-адреналовой системы в борьбе за существование первостепенно. У человека она также играет важную роль при подготовке организма к выполнению повышенных нагрузок, например при подготовке спортсменов к соревнованию,
В действии адреналина и норадреналина есть некоторые различия. Так, норадреналин в отличие от адреналина лишь в малой степени увеличивает содержание глюкозы в крови и поглощение О2.
Корковый слой надпочечников в нормальных условиях секретирует адренокортикостероиды: кортизол (гидрокортизон), кортикостерон и альдостерон. Предшественником адренокортикостероидов является холестерин.
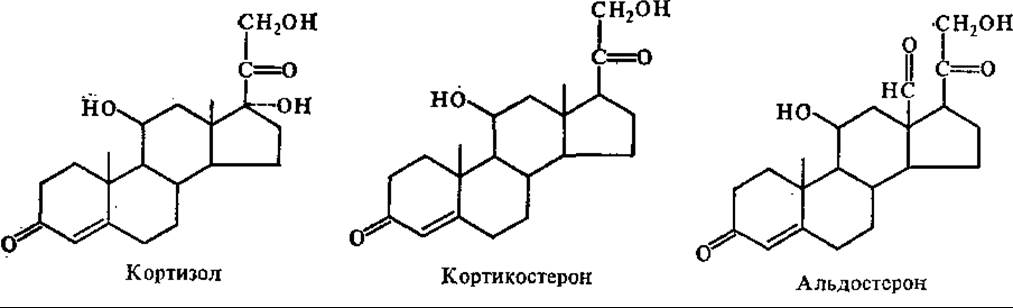
У человека кортизол секретируется в количестве 10—30 мг/сут, кортикостерон — 2—4 мг, альдостерон — 300—400 мкг. Кортизол и кортикостерон транспортируются сиглобулином — транскортином, а альдостерон — альбумином. Кортикостероиды оказывают влияние на следующие метаболические и физиологические процессы: 1) способствуют гликогенезу, повышают содержание гликогена в печени и глюкозы в крови; 2) сильно тормозят синтез белков в мышцах и ряде других тканей, но активируют образование специфических белков печени; 3) усиливают мобилизацию липидов из запасных отложений; 4) регулируют водно-солевой обмен; 5) стимулируют эритропоэз, но уменьшают количество лимфоцитов; 6) усиливают секрецию НСl и пепсиногена; 7) предотвращают и замедляют развитие воспалительных, аллергических явлений; 8) способствуют поддержанию гомеостаза при действии повреждающих факторов (травмы, инфекционные агенты, вредные химикаты и т. д.).
12.6.7. Поджелудочная железа. Синтезирует инсулин, глюкагон (см. разд. 6.8), соматостатин, секретин, птичий панкреатический полипептид (АРР, от лат. avis — птица).
Соматостатин ингибирует высвобождение соматотропина гипофизом, инсулина и глюкагона поджелудочной железой, гастрина слизистой желудка. В связи с этим соматостатин регулирует секрецию гормона роста, оказывает антигипергликемическое действие, способствует высвобождению липидов из жировой ткани. Присутствие соматостатина показано не только в гипоталамусе — основном месте его секреции, но и в поджелудочной железе, слизистой желудка, кишечника. По строению соматостатин представляет собой четырнадцатичленный пептид с одной дисульфидной связью.
Секретин — содержит 27 аминокислотных остатков, стимулирует выделение панкреатического сока и в меньшей степени желчи, кишечного сока, угнетает образование гастрина. У животных и человека помимо гастрина и секретина вырабатываются и другие гормоны пептидной природы, регулирующие деятельность желудочно-кишечного тракта — холецистокинин (стимулирует сокращение желчного пузыря), сосудисто-активный кишечный пептид, желудочный ингибирующий пептид и др.
Панкреатический полипептид содержит 36 аминокислот. Увеличивает секрецию желудочных и панкреатических ферментов, расслабляет желчный пузырь, уменьшает перистальтику кишечника.
12.6.8. Тимус. Осуществляет синтез и секрецию гормонов, которые влияют на скорость развития и созревания определенных популяций лимфоидных клеток. К таким гормонам относятся: тимозин, тимопоэтины I и II, тимусный гуморальный фактор и гомеостатический тимусный гормон. Поскольку лимфоидные клетки имеют первостепенное значение и для иммунитета, роль гормонов тимуса в этом плане тоже очень важна.
12.6.9. Гипофиз. Придаток мозга, состоящий из передней (аденогипофиз), задней (нейрогипофиз) и промежуточной долей. Международная комиссия по анатомической номенклатуре подразделяет гипофиз на нейрогипофиз и аденогипофиз, включая в последний переднюю долю и промежуточную (среднюю).
Нейрогипофиз млекопитающих секретирует: вазопрессин, окситоцин, а- и ß-меланоцитстимулирующие гормоны, когерин.
Окситоцин и вазопрессин — циклические нонапептиды. Поскольку два остатка цистеина, замыкая пептид в цикл, образуют одну молекулу цистеина, эти гормоны иногда называют октапептидами.
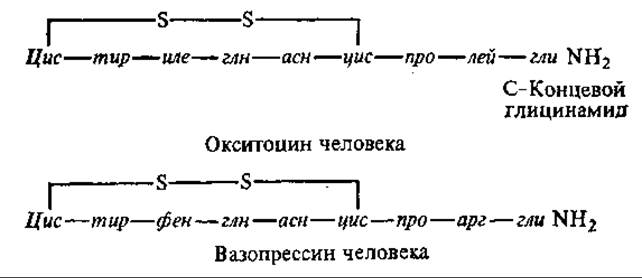
Вазопрессин и окситоцин человека отличаются лишь двумя аминокислотными остатками. Изучение структуры вазопрессина у ряда высших животных показало наличие видовой специфичности в его строении. Так, вазопрессин свиньи отличается от вазопрессина человека заменой остатка арг на лиз.
Окситоцин стимулирует сокращение мышц матки, активирует лактацию. Вазопрессин усиливает сокращение гладких мышц кровеносных сосудов, что приводит к повышению давления. Кроме того, вазопрессин участвует в регуляции осмотического давления плазмы крови, водного обмена (обладает антидиуретическим свойством, т. е. препятствует выделению мочи).
Когерин — пептид, синтезирующийся в нейрогипофизе и вызывающий координированное сокращение кишечника.
Меланоцитстимулирующий гормон (меланотропин, МСГ) обладает способностью увеличивать число меланоцитов — клеток, несущих темный пигмент — меланин. У позвоночных животных этой способностью обладают два пептида: а-МСГ и ß-МСГ. Их аминокислотный состав у отдельных видов позвоночных имеет некоторые отличия.
Хорошо доказана способность МСГ вызывать потемнение кожи земноводных за счет диспергирования меланина, концентрированного в центре меланоцитов. У человека и высших животных МСГ повышает активность щитовидной железы, стимулирует секрецию сальных желез, снижает адаптацию глаз к темноте, повышает чувствительность к свету, участвуя в регуляции движения клеток черного пигментного слоя в глазу.
Нейрогоморны появились на очень ранних этапах филогенеза. У всех позвоночных, кроме млекопитающих, секретируется вазотоцин, отличающийся от вазопрессина только заменой остатка фен на иле. Этот гормон вырабатывается даже у древних рыб и круглоротых. Окситоцин также является продуктом длительной эволюции. У древних рыб аналогом этого гормона является глумитоцин (отличается от окситоцина двумя аминокислотными остатками в 4-м и 8-м положении), у костистых рыб изотоцин (тоже отличия по двум аминокислотам), а у птиц, рептилий и амфибий — мезотоцин (отличается от окситоцина только заменой остатка лей в 8-м положении на иле).
В процессе эволюции произошли и изменения в функциональном значении гормонов нейрогипофиза. Так вазотоцин, экстрагированный из гипофиза рыб, оказывает сильное антидиуретическое действие при введении его амфибиям и млекопитающим, но не влияет таким путем на рыб. Видимо, у последних вазотоцин не регулирует водный обмен, а выполняет какую-то иную роль.
Аденогипофиз секретирует тиреотропин, адренокортикотропин, лютеинизирующий и фолликулостимулирующий гормоны, пролактин, соматотропин, липотропины.
Тиреотропин — гликопротеин, состоящий из двух субъединиц (М13 600 и 14 700); относится к числу белков, богатых серой. Влияет на скорость поглощения иода из крови в щитовидной железе, включение иода в состав тиреоидных гормонов и на секрецию последних.
Адренокортикотропный гормон (адренокортикотропин, кортикотропин, АКТГ) представляет собой пептид, образованный 39 остатками аминокислот. Первичная структура АКТГ установлена для человека и нескольких представителей высших животных (овца, свинья, бык и др.). Обнаружены видовые различия: наиболее вариабельными являются аминокислоты в положениях 25—32. АКТГ участвует в регуляции биосинтеза кортикостероидов надпочечниками, стимулирует рост коры надпочечников, участвует в процессе мобилизации липидов из жировой ткани.
Фолликулостимулирующий, лютеинизирующий гормоны и пролактин объединяют под названием гонадотропные гормоны. Это гликопротеины, состоящие из а и ß-субъединиц. Фолликулостимулирующий гормон вызывает у самок развитие большого числа фолликулов и увеличение массы яичников. Лютеинизирующий гормон — окончательное созревание фолликулов яичника, разрыв фолликула и превращение его в желтое тело. Пролактин необходим для появления молока у самок млекопитающих при родах.
Соматотропин — гормон, ускоряющий рост и увеличивающий массу тела. Это белок с молекулярной массой 21 000.
Липотропины а и ß-полипептиды, стимулирующие высвобождение жирных кислот из жировой ткани. ß-Липотропин состоит из 91 аминокислотного остатка. Является предшественником эндорфинов (см. разд. 2.5.1). Кроме того, цепь аминокислотных остатков ß-ли- потропина содержит последовательности АКТГ и ß-меланоцитстимулирующего гормона.
12.6.10. Другие гормоны млекопитающих. Некоторые гормоны синтезируются в клетках, не относящихся к эндокринной системе. Так, слизистой желудка вырабатывается группа гормонов, получивших название гастрини. Основная их функция — стимуляция секреции желудочного сока, соляной кислоты. Кроме того, они усиливают тонус и сокращение мышц желудка и тонкого кишечника, стимулируют секрецию глюкагона и инсулина поджелудочной железой. Показано наличие в слизистой желудка четырех гастринов, один из них обладает молекулярной массой, равной 20 000, и относится к белкам, а остальные три являются пептидами.
12.6.11. Гормоны насекомых. Беспозвоночные обладают нейроэндокринной системой, которая имеет ряд общих черт с нейроэндокринной системой млекопитающих. Насекомые синтезируют гормон активации, который регулирует секреторную функцию парных желез. Парные железы вырабатывают ювенильный гормон у личинок насекомых.
Проторокальная железа насекомых секретирует экдизон. Он относится к стероидным гормонам. Этот гормон необходим для периодической смены экзоскелета личинок, в связи с чем и получил дополнительное название гормона линьки. Обнаружены различные виды экдизонов, считают, что они могут оказывать влияние на разные стадии развития насекомых. Экдизон стимулирует синтез РНК в тканях, по-видимому, подобно стероидным гормонам млекопитающих, непосредственно контролирует процесс транскрипции.