БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ - Е. С. Северина - 2004
РАЗДЕЛ 2. ЭНЗИМОЛОГИЯ
V. Основы кинетики ферментативных реакций
Кинетика ферментативных реакций — раздел энзимологии, изучающий зависимость скорости химических реакций, катализируемых ферментами, от химической природы реагирующих веществ, а также от факторов окружающей среды.
Для измерения каталитической активности ферментов используют такие показатели, как скорость реакции или активность фермента. Скорость ферментативной реакции определяется изменением количества молекул субстрата или продукта за единицу времени. Скорость ферментативной реакции — мера каталитической активности фермента, её обозначают как активность фермента.
Математически скорость ферментативной реакции выражается в изменении концентрации субстрата (уменьшение) или продукта (увеличение) за единицу времени:
V = D[S]/t = D[Р]/t.
На начальном этапе [0 — t0] скорость реакции прямо пропорциональна времени и имеет линейную зависимость. Графически изменение скорости ферментативной реакции определяется тангенсом угла наклона касательной к кривой профиля реакции. Чем больше угол наклона, тем больше изменение скорости реакции (рис. 2-15).
Рис. 2-15. Зависимость накопления продукта (А) и убыли субстрата (Б) от времени (продолжительности) протекания реакции. Скорость ферментативной реакции определяется изменением концентрации продукта или субстрата за единицу времени. В реакциях, катализируемых ферментами 1 и 2, начальная скорость реакции, катализируемой ферментом 1, ниже, чем скорость реакции, катализируемой ферментом 2, так как тангенс угла наклона касательной к кривой профиля реакции, проведённой из «0» точки у второго фермента выше, как в случае накопления продукта (А), так и убыли субстрата (Б). Скорость в любой момент времени t определяется тангенсом угла наклона касательной к профилю реакции в момент времени t. Период времени ферментативной реакции [t0 - t1] характеризуется линейным накоплением продукта (или убылью субстрата) в зависимости от длительности реакции. Период ферментативной реакции [t1 - tx] характеризуется нелинейным накоплением продукта (или убылью субстрата) в зависимости от времени реакции.

С течением времени изменение скорости ферментативной реакции в экспериментальных условиях уменьшается, об этом свидетельствует уменьшение угла наклона касательной в момент времени 1. Снижение скорости ферментативной реакции может происходить за счёт ряда факторов: уменьшения концентрации субстрата, увеличения концентрации продукта, который может оказывать ингибирующее действие, могут происходить изменения pH раствора, инактивация фермента и т. д.
На этапе [t1- tx] скорость реакции изменяется нелинейно в зависимости от времени. Поэтому для определения скорости ферментативной реакции чаще всего исследуют изменение скорости на начальном этапе [t0 - t1], где наблюдают линейное изменение концентрации продукта (или субстрата).
Скорость ферментативной реакции зависит от ряда факторов, таких как количество и активность ферментов, концентрация субстрата, температура среды, pH раствора, присутствие регуляторных молекул (активаторов и ингибиторов). Рассмотрим влияние этих факторов на скорость ферментативной реакции.
А. Зависимость скорости ферментативной реакции от количества ферментов
При проведении ферментативной реакции в условиях избытка субстрата скорость реакции будет зависеть от концентрации фермента. Графическая зависимость такой реакции имеет вид прямой линии (рис. 2-16). Однако количество фермента часто невозможно определить в абсолютных величинах, поэтому на практике пользуются условными величинами, характеризующими активность фермента: одна международная единица активности (МЕ) соответствует такому количеству фермента, которое катализирует превращение 1 мкмоль субстрата за 1 мин при оптимальных условиях проведения ферментативной реакции. Оптимальные условия индивидуальны для каждого фермента и зависят от температуры среды, pH раствора, при отсутствии активаторов и ингибиторов.

Рис. 2-16. Зависимость скорости ферментативной реакции (V) от концентрации фермента.

Количество единиц активности nМЕ определяют по формуле:

В 1973 г. была принята новая единица активности ферментов: 1 катал (кат), соответствующий такому количеству катализатора, которое превращает 1 моль субстрата за 1 с. Количество каталов определяют по формуле:

Международная единица ферментативной активности МЕ связана с каталом следующими равенствами:
1 кат = 1 моль S/с = 60 моль S/мин = 60 х 106 мкмоль/мин = 6 х 107 МЕ,
1 МЕ = 1 мкмоль/мин = 1/60 мкмоль/с = 1/60 мккат = 16,67 нкат.
В медицинской и фармацевтической практике для оценки активности ферментов часто используют международные единицы активности — МЕ. Для оценки количества молекул фермента среди других белков данной ткани определяют удельную активность (уд. ак.) фермента, численно равную количеству единиц активности фермента (nМЕ) в образце ткани, делённому на массу (мг) белка в этой ткани:
![]()
По удельной активности судят об очистке фермента: чем меньше посторонних белков, тем выше удельная активность.
Б. Зависимость скорости ферментативной реакции от температуры среды
Повышение температуры до определённых пределов оказывает влияние на скорость ферментативной реакции, подобно влиянию температуры на любую химическую реакцию. С повышением температуры ускоряется движение молекул, что приводит к повышению вероятности взаимодействия реагирующих веществ. Кроме того, температура может повышать энергию реагирующих молекул, что также приводит к ускорению реакции. Однако скорость химической реакции, катализируемая ферментами, имеет свой температурный оптимум, превышение которого сопровождается понижением ферментативной активности, возникающим из-за термической денатурации белковой молекулы (рис. 2-17).
Рис. 2-17. Зависимость скорости ферментативной реакции (V) от температуры.
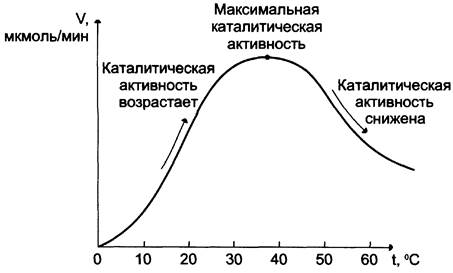
Для большинства ферментов человека оптимальна температура 37— 38 °С. Однако в природе существуют и термостабильные ферменты. Например, Taq — полимераза, выделенная из микроорганизмов, живущих в горячих источниках, не инактивируется при повышении температуры до 95 °С. Этот фермент используют в научно- практической медицине для молекулярной диагностики заболеваний с использованием метода полимеразной цепной реакции (ПЦР).
В. Зависимость скорости ферментативной реакции от рН среды
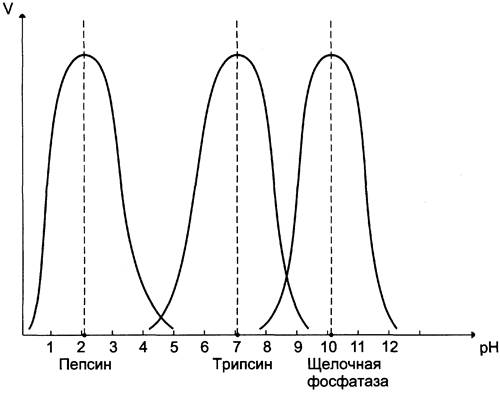
Активность ферментов зависит от pH раствора, в котором протекает ферментативная реакция. Для каждого фермента существует значение pH, при котором наблюдается его максимальная активность. Отклонение от оптимального значения pH приводит к понижению ферментативной активности.
Влияние pH на активность ферментов связано с ионизацией функциональных групп аминокислотных остатков данного белка, обеспечивающих оптимальную конформацию активного центра фермента. При изменении pH от оптимальных значений происходит изменение ионизации функциональных групп молекулы белка. Например, при закислении среды происходит протонирование свободных аминогрупп (NH3+), а при защелачивании происходит отщепление протона от карбоксильных групп (СОО-). Это приводит к изменению конформации молекулы фермента и конформации активного центра; следовательно, нарушается присоединение субстрата, кофакторов и коферментов к активному центру. Кроме того, pH среды может влиять на степень ионизации или пространственную организацию субстрата, что также влияет на сродство субстрата к активному центру. При значительном отклонении от оптимального значения pH может происходить денатурация белковой молекулы с полной потерей ферментативной активности.
Оптимум значения pH у разных ферментов различный (рис. 2-18). Ферменты, работающие в кислых условиях среды (например, пепсин в желудке или лизосомальные ферменты), эволюционно приобретают конформацию, обеспечивающую работу фермента при кислых значениях pH. Однако большая часть ферментов организма человека имеет оптимум pH, близкий к нейтральному, совпадающий с физиологическим значением pH (табл. 2-1).
Таблица 2-1. Оптимальные значения pH для некоторых ферментов
Фермент |
Оптимальное значение pH |
Пепсин |
1,5-2 |
Пируваткарбоксилаза |
4,8 |
Каталаза |
6,8-7 |
Фумараза |
6,5 |
Уреаза |
6,8-7,2 |
Кабоксипептидаза |
7,5 |
Трипсин |
6,5-7,5 |
Аргиназа |
9,5-9,9 |
Рис. 2-18. Зависимость скорости ферментативной реакции (V) от pH среды.
Г. Зависимость скорости ферментативной реакции от количества субстрата
Если концентрацию ферментов оставить постоянной, изменяя только количество субстрата, то график скорости ферментативной реакции описывают гиперболой (рис. 2-19).
Рис. 2-19. Зависимость скорости реакции (V) от концентрации субстрата S. \/mах — максимальная скорость реакции при данной концентрации фермента в оптимальных условиях проведения реакции. Кm — константа Михаэлиса.

При увеличении количества субстрата начальная скорость возрастает. Когда фермент становится полностью насыщенным субстратом, т. е. происходит максимально возможное при данной концентрации фермента формирование фермент-субстратного комплекса, наблюдают наибольшую скорость образования продукта. Дальнейшее повышение концентрации субстрата не приводит к увеличению образования продукта, т. е. скорость реакции не возрастает. Данное состояние соответствует максимальной скорости реакции Vmах.
Таким образом, концентрация фермента — лимитирующий фактор в образовании продукта. Это наблюдение легло в основу ферментативной кинетики, разработанной учёными Л. Михаэлисом и М. Ментен в 1913 г.
Ферментативный процесс можно выразить следующим уравнением:

где к1 — константа скорости образования фермент-субстратного комплекса; к-1 — константа скорости обратной реакции, распада фермент- субстратного комплекса; к2 — константа скорости образования продукта реакции.
Следующее соотношение констант скоростей (к-1 + к2)/к1 называют константой Михаэлиса и обозначают Кm.
Скорость реакции пропорциональна концентрации фермент-субстратного комплекса ЕS, а скорость образования ЕS зависит от концентрации субстрата и концентрации свободного фермента. На концентрацию ЕS влияет скорость формирования и распада ЕS.
Наибольшая скорость реакции наблюдается в том случае, когда все молекулы фермента находятся в комплексе с субстратом, т. е. в фермент- субстратном комплексе ЕS, т.е. [Е] = [ЕS].
Зависимость скорости ферментативной реакции от концентрации субстрата выражается следующим уравнением (математическое выведение этой формулы можно найти в пособиях по ферментативной кинетике):
![]()
Это уравнение получило название уравнения Михаэлиса — Ментен.
В случае, когда скорость реакции равна половине максимальной, Кm = [S] (рис. 2-19). Таким образом, константа Михаэлиса численно равна концентрации субстрата, при которой достигается половина максимальной скорости.
Уравнение Михаэлиса — Ментен — основное уравнение ферментативной кинетики, описывающее зависимость скорости ферментативной реакции от концентрации субстрата.
Если концентрация субстрата значительно больше Кm (S» Кm), то увеличение концентрации субстрата на величину Кm практически не влияет на сумму (Кm + S) и её можно считать равной концентрации субстрата. Следовательно, скорость реакции становится равной максимальной скорости: V = Vmах. В этих условиях реакция имеет нулевой порядок, т. е. не зависит от концентрации субстрата. Можно сделать вывод, что Vmах — величина постоянная для данной концентрации фермента, не зависящая от концентрации субстрата.
Если концентрация субстрата значительно меньше Кm (S «Кm), то сумма (Кm + S) примерно равна Кm, следовательно, V = Vmах[S]/Кm, т. е. в данном случае скорость реакции прямо пропорциональна концентрации субстрата (реакция имеет первый порядок).
Vmах и Кm — кинетические характеристики эффективности фермента. Vmах даёт характеристику каталитической активности фермента и имеет размерность скорости ферментативной реакции моль/л, т. е. определяет максимальную возможность образования продукта при данной концентрации фермента и в условиях избытка субстрата.
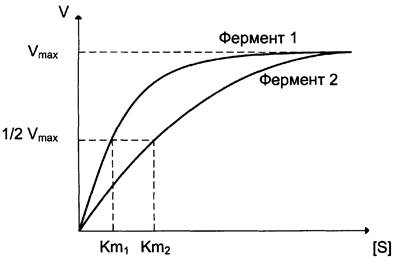
Кm характеризует сродство данного фермента к данному субстрату и является величиной постоянной, не зависящей от концентрации фермента. Чем меньше Кm, тем больше сродство фермента к данному субстрату, тем выше начальная скорость реакции и наоборот, чем больше Кm, тем меньше начальная скорость реакции, тем меньше сродство фермента к субстрату.
На рис. 2-20 представлена зависимость скорости двух ферментативных реакций (1 и 2) от концентрации субстрата. Константа Михаэлиса первого фермента меньше константы Михаэлиса второго фермента (Кm1 <Кm2). Следовательно, сродство первого фермента к субстрату выше, чем у второго фермента, и начальная скорость реакции, катализируемой первым ферментом, выше в сравнении со вторым ферментом.
Рис. 2-20. Влияние различных концентраций субстрата на скорость реакции, катализируемой ферментами 1 и 2.