БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ - Е. С. Северина - 2004
РАЗДЕЛ 5. БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Все клетки имеют мембраны. Кроме того, почти во всех эукариотических клетках существуют органеллы, каждая из которых имеет свою мембрану. Мембраны ответственны за выполнение многих важнейших функций клетки. Согласованное функционирование мембранных систем — рецепторов, ферментов, транспортных механизмов помогает поддерживать гомеостаз клетки и в то же время быстро реагировать на изменения внешней среды.
К основным функциям мембран можно отнести:
✵ отделение клетки от окружающей среды и формирование внутриклеточных компартментов (отсеков);
✵ контроль и регулирование транспорта огромного разнообразия веществ через мембраны;
✵ участие в обеспечении межклеточных взаимодействий, передаче внутрь клетки сигналов;
✵ преобразование энергии пищевых органических веществ в энергию химических связей молекул АТФ.
I. Роль мембран в метаболизме и их разнообразие
Основные принципы структурной организации всех мембран одинаковы, однако одна из самых характерных особенностей — огромное их разнообразие. Мембраны органелл эукариотических клеток уникальны по своему составу и по характеру выполняемых функций.
Плазматическая мембрана
Плазматическая мембрана, окружающая каждую клетку, определяет её величину, обеспечивает транспорт малых и больших молекул из клетки и в клетку, поддерживает разницу концентраций ионов по обе стороны мембраны. Мембрана участвует в межклеточных контактах, воспринимает, усиливает и передаёт внутрь клетки сигналы внешней среды. С мембраной связаны многие ферменты, катализирующие биохимические реакции.
Ядерная мембрана
Ядерная оболочка состоит из внешней и внутренней ядерных мембран. Ядерная оболочка имеет поры, через которые РНК проникают из ядра в цитоплазму, а регуляторные белки из цитоплазмы в ядро.
Внутренняя ядерная мембрана содержит специфические белки, имеющие участки связывания основных полипептидов ядерного матрикса — ламина А, ламина В и ламина С. Важная функция этих белков — дезинтеграция ядерной оболочки в процессе митоза.
Мембрана эндоплазматического ретикулума (ЭР)
Мембрана ЭР имеет многочисленные складки и изгибы. Она образует непрерывную поверхность, ограничивающую внутреннее пространство, называемое полостью ЭР. Шероховатый ЭР связан с рибосомами, на которых происходит синтез белков плазматической мембраны, ЭР, аппарата Гольджи, лизосом, а также секретируемых белков. Области ЭР, не содержащие рибосом, называют гладким ЭР. Здесь происходит завершающий этап биосинтеза холестерина, фосфолипидов, реакции окисления собственных метаболитов и чужеродных веществ с участием мембранных ферментов — цитохрома Р450, цитохром Р450 редуктазы, цитохром b5 редуктазы и цитохрома b5 (см. раздел 12).
Аппарат Гольджи
Аппарат Гольджи — важная мембранная органелла, отвечающая за модификацию, накопление, сортировку и направление различных веществ в соответствующие внутриклеточные компартменты, а также за пределы клетки. Специфические ферменты мембраны комплекса Гольджи, гликозилтрансферазы, гликозилируя белки по остаткам серина, треонина или амидной группе аспарагина, завершают образование сложных белков — гликопротеинов.
Митохондриальные мембраны
Митохондрии — органеллы, окружённые двойной мембраной, специализирующиеся на синтезе АТФ путём окислительного фосфорилирования. Отличительная особенность внешней митохондриальной мембраны — содержание большого количества белка порина, образующего поры в мембране. Благодаря порину внешняя мембрана свободно проницаема для неорганических ионов, метаболитов и даже небольших молекул белков (меньше 10 кД). Для больших белков внешняя мембрана непроницаема, это позволяет митохондриям удерживать белки межмембранного пространства от утечки в цитозоль.
Для внутренней мембраны митохондрий характерно высокое содержание белков, около 70%, которые выполняют в основном каталитическую и транспортную функции. Транслоказы мембраны обеспечивают избирательный перенос веществ из межмембранного пространства в матрикс и в обратном направлении, ферменты участвуют в транспорте электронов (цепи переноса электронов) и синтезе АТФ. Подробно строение и функционирование ферментов цепи переноса электронов рассмотрено в разделе 6.
Мембрана лизосом
Мембрана лизосом играет роль «щита» между активными ферментами (более 50), обеспечивающими реакции распада белков, углеводов, жиров, нуклеиновых кислот, и остальным клеточным содержимым. Мембрана содержит уникальные белки, например, АТФ-зависимую протонную помпу (насос), которая поддерживает кислую среду (pH 5), необходимую для действия гидролитических ферментов (протеаз, липаз), а также транспортные белки, позволяющие продуктам расщепления макромолекул покидать лизосому. Большинство белков лизосомальной мембраны сильно гликозилированы, углеводные составляющие, находящиеся на внутренней поверхности мембраны, защищают их от действия протеаз.
А. Строение и состав мембран
Биологические мембраны представляют собой «ансамбли» липидных и белковых молекул, удерживаемых вместе с помощью нековалентных взаимодействий.
Основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолипиды и гликолипиды. Липидный бислой образован двумя рядами липидов, гидрофобные радикалы которых спрятаны внутрь, а гидрофильные группы обращены наружу и контактируют с водной средой. Белковые молекулы как бы «растворены» в липидном бислое (рис. 5-1).
Рис. 5-1. Поперечный разрез плазматической мембраны.

1. Структура и свойства липидов мембран
Мембранные липиды — амфифильные (амфипатические) молекулы, т. е. в молекуле есть как гидрофильные группы (полярные «головки»), так и алифатические радикалы (гидрофобные «хвосты»), самопроизвольно формирующие бислой. В большинстве эукариотических клеток они составляют около 30-70% массы мембраны (рис. 5-2). В мембранах присутствуют липиды трёх главных типов — фосфолипиды, гликолипиды и холестерол (холестерин).
Рис. 5-2. Содержание липидов и белков в различных клеточных мембранах (%).
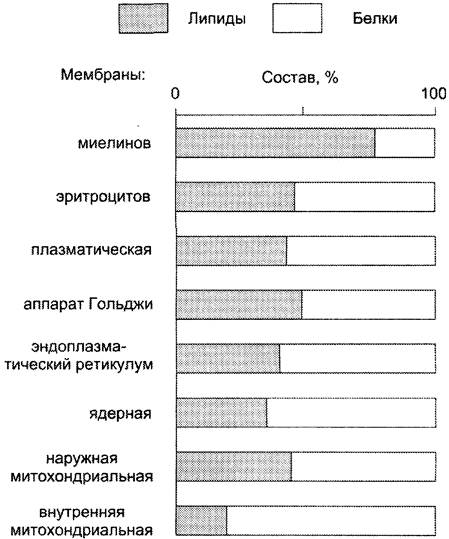
Липидный состав мембран различен, содержание того или другого липида, по-видимому, определяется разнообразием функций, выполняемых этими липидами в мембранах.
Фосфолипиды. Все фосфолипиды можно разделить на 2 группы — глицерофосфолипиды и сфингофосфолипиды. Глицерофосфолипиды относят к производным фосфатидной кислоты. Наиболее распространённые глицерофосфолипиды мембран — фосфатидилхолины и фосфатидилэтаноламины (рис. 5-3). В мембранах эукариотических клеток обнаружено огромное количество разных фосфолипидов, причём они распределены неравномерно по разным клеточным мембранам. Эта неравномерность относится к распределению как полярных «головок» (табл. 5-1), так и ацильных остатков (табл. 5-2).
Таблица 5-1. Фосфолипидный состав клеточных органелл и плазматической мембраны гепатоцитов
Фосфолипиды с разным строением полярных «головок» |
Доля от суммарного количества фосфолипидов, % |
||||
митохондрии |
лизосомы |
Ядерная мембрана |
мембраны аппарата Г ольджи |
Плазматическая мембрана |
|
Кардиолипин |
18 |
1 |
4 |
1 |
1 |
Фосфатидилэтаноламин |
35 |
14 |
13 |
20 |
23 |
Фосфатидилхолин |
40 |
40 |
55 |
50 |
39 |
Фосфатидилинозитол |
5 |
5 |
10 |
12 |
8 |
Фосфатидилсерин |
1 |
2 |
3 |
6 |
9 |
Фосфатидная кислота |
— |
1 |
2 |
1 |
1 |
Сфингомиелин |
1 |
20 |
3 |
8 |
16 |
Таблица 5-2. Жирно-кислотный состав некоторых мембран печени
Жирные кислоты, % (по массе) |
Мембранная фракция |
||||
Мембраны митохондрий |
ЭР |
Аппарат Гольджи |
Плазматическая мембрана |
||
наружная |
внутренняя |
||||
Миристиновая 14:0 |
0,4 |
0,3 |
0,4 |
0,9 |
0,9 |
Пальмитиновая 16:0 |
4,0 |
3,6 |
3,1 |
— |
— |
Пальмитоолеиновая 16:1 |
21,0 |
18,0 |
26,5 |
22,5 |
31.2 |
Стеариновая 18:0 |
13,5 |
15,8 |
14,9 |
18,5 |
12,9 |
Арахидоновая 20:4 |
15,7 |
18,5 |
14,0 |
14,5 |
11,1 |
Цервоновая 22:6 |
3,5 |
3,8 |
0,7 |
— |
— |
Каждый глицерофосфолипид, например, фосфатидилхолин, представлен несколькими десятками фосфатидилхолинов, отличающихся друг от друга строением жирно-кислотных остатков.
На долю глицерофосфолипидов (полярная группа — инозитол) приходится лишь 2 — 8% всех фосфолипидов, содержащихся в клеточной мембране эукариотов. Инозитол в составе фосфатидилинозитолов может быть фосфорилирован по С4 (фосфатидилинозитол-4-монофосфат) или С4 и С5 (фосфатидилинозитол-4,5-бисфосфат).
Рис. 5-3. Глицерофосфолипиды мембран.

В состав фосфатидилинозитол-4,5-бисфосфатов входят в основном ацильные остатки стеариновой или пальмитиновой (по первому положению глицерола) и арахидоновой (по второму положению) жирных кислот.
Специфические фосфолипиды внутренней мембраны митохондрий — кардиолипины (дифосфатидилглицеролы), построенные на основе глицерола и двух остатков фосфатидной кислоты. Они синтезируются ферментами внутренней мембраны митохондрий и составляют около 22% от всех фосфолипидов мембраны.
В плазматических мембранах клеток в значительных количествах содержатся сфингомиелины (рис. 5-4). Сфингомиелины построены на основе церамида — ацилированного аминоспирта сфингозина. Полярная группа состоит из остатка фосфорной кислоты и холина, этаноламина или серина. Сфингомиелины — главные липиды миелиновой оболочки нервных волокон.
Рис. 5-4. Сфингофосфолипиды — производные церамида.

Гликолипиды. В гликолипидах гидрофобная часть представлена церамидом. Гидрофильная группа — углеводный остаток, присоединённый гликозидной связью к гидроксильной группе у первого углеродного атома церамида (рис. 5-5). В зависимости от длины и строения углеводной части различают цереброзиды, содержащие моно- или олигосахаридный остаток, и ганглиозиды, к ОН-группе которых присоединён сложный, разветвлённый олигосахарид, содержащий N-ацетилнейраминовую кислоту (NANA) (см. раздел 7).
Рис. 5-5. Гликолипиды. Gal — галактоза; Glc— глюкоза; NANA (NeuAc) — N-ацетилнейраминовая или опаловая кислота.
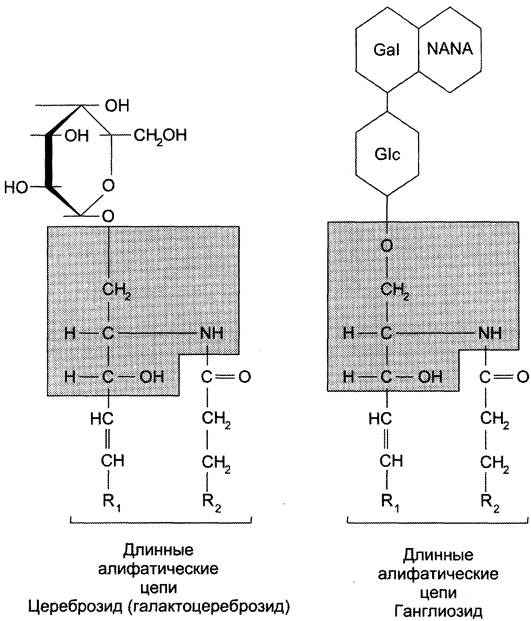
Полярные «головки» гликосфинголипидов находятся на наружной поверхности плазматических мембран. В значительных количествах гликолипиды содержатся в мембранах клеток мозга, эритроцитов, эпителиальных клеток. Ганглиозиды эритроцитов разных индивидуумов различаются строением олигосахаридных цепей, проявляющих антигенные свойства.
Холестерол. Холестерол присутствует во всех мембранах животных клеток. Его молекула состоит из жёсткого гидрофобного ядра и гибкой углеводородной цепи, единственная гидроксильная группа является «полярной головкой» (рис. 5-6).
Рис. 5-6. Положение молекулы холестерола в мембране. Молекула холестерола располагается в липидном слое мембраны параллельно алифатическим цепям молекул фосфо- и гликолипидов. Гидроксильная группа холестерола контактирует с гидрофильными «головками» этих липидов.
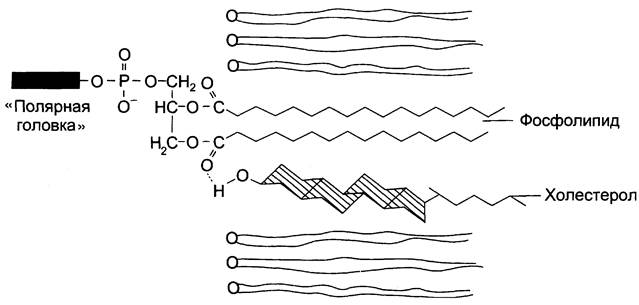
Для животной клетки среднее молярное отношение холестерол/фосфолипиды равно 0,3 — 0,4, но в плазматической мембране это соотношение гораздо выше (0,8 — 0,9). Наличие холестерола в мембранах уменьшает подвижность жирных кислот, снижает латеральную диффузию липидов и белков, и поэтому может влиять на функции мембранных белков.
В составе мембран растений холестерола нет, а присутствуют растительные стероиды — ситостерол и стигмастерол.
2. Трансмембранная асимметрия липидов
Каждая мембрана клетки замкнута, т. е. имеет внутреннюю и внешнюю поверхности, различающиеся по липидному и белковому составам — эту особенность мембран называют трансмембранной (поперечной) асимметрией.
Липидная асимметрия возникает прежде всего потому, что липиды с более объёмными полярными «головками» стремятся находиться в наружном монослое, так как там площадь поверхности, приходящаяся на полярную «головку», больше. Фосфатидилхолины и сфингомиелины локализованы преимущественно в наружном монослое, а фосфатидилэтаноламины и фосфатидилсерины в основном во внутреннем.
Липиды в некоторых биологических мембранах с довольно большой частотой мигрируют с одной стороны мембраны на другую, т. е. совершают «флип-флоп» (от англ. flip-flop) перескоки (рис. 5 — 7). Перемещение липидных молекул затрудняют полярные «головки», поэтому липиды, находящиеся на внутренней стороне мембраны, имеют относительно высокую скорость трансмембранной миграции по сравнению с липидами наружной стороны мембраны, мигрирующих медленнее или вообще не совершающими «флип-флоп» перескоки.
Рис. 5-7. Типы движений липидных молекул в бислое мембран.
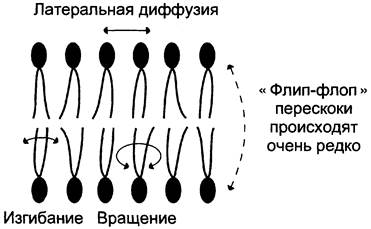
3. Жидкостность мембран
Для мембран характерна жидкостность (текучесть), способность липидов и белков к латеральной диффузии. Скорость перемещения молекул зависит от микровязкости мембран, которая, в свою очередь, определяется относительным содержанием насыщенных и ненасыщенных жирных кислот в составе липидов. Микровязкость меньше, если в составе липидов преобладают ненасыщенные жирные кислоты, и больше при высоком содержании насыщенных жирных кислот.
Ацильные (алифатические) остатки ненасыщенных жирных кислот имеют так называемые «изломы» (см. раздел 8). Эти «изломы» препятствуют слишком плотной упаковке молекул в мембране и делают её более рыхлой, а, следовательно, и более «текучей». На текучесть мембран также влияют размеры углеводородных «хвостов» липидов, с увеличением длины которых мембрана становится более «текучей».
4. Функции мембранных липидов
Фосфо- и гликолипиды мембран, помимо участия в формировании липидного бислоя, выполняют ряд других важных функций.
Липиды формируют среду для функционирования мембранных белков, принимающих в ней нативную конформацию. Выделенные из мембран ферменты, лишённые липидного окружения, как правило, не проявляют каталитической активности.
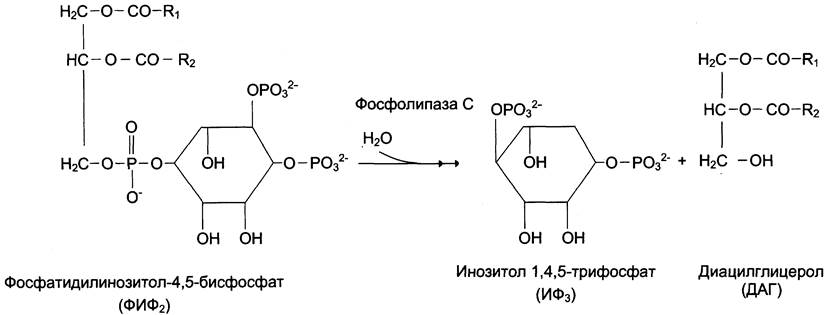
Некоторые мембранные липиды — предшественники вторичных посредников при передаче гормонального сигнала. Так, фосфати- дилинозитол-4,5-бисфосфат (ФИФ2) под действием фермента фосфолипазы С гидролизуется до диацилглицерола (ДАГ), активатора про- теинкиназы С и инозитол-1,4,5-трифосфата (ИФ3) — регулятора кальциевого обмена в клетке (рис. 5-8). ДАГ, ИФ3, протеинкиназа С и Са2+ — участники инозитолфосфатной системы передачи сигнала.
Рис. 5-8. Гидролиз фосфатидилинозитол-4,5-бисфосфата.
Кроме того, некоторые липиды выполняют «якорную» функцию, например, к фосфатидил- инозитолам через олигосахарид могут присоединяться специфические белки наружной поверхности клетки (рис. 5-9). Фосфатидилинозитол с присоединённым к нему олигосахаридом (гликаном) называют фосфатидилинозитолгликаном.
Рис. 5-9. «Якорная» функция фосфатидилинозитолгликанов.
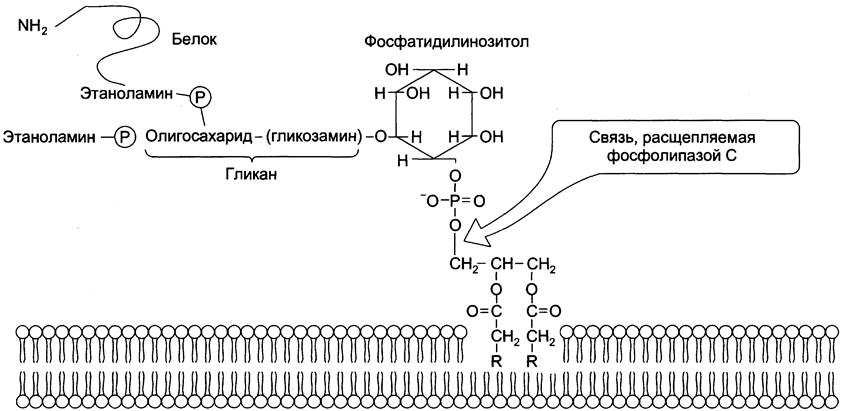
Связь белков с этой молекулой (гликаном) осуществляется через фосфоэтаноламин. Пример такого «заякоренного» белка — ацетилхолинэстераза, катализирующая гидролиз ацетилхолина в синаптической щели. Этот фермент фиксируется на постсинаптической мембране, ковалентно присоединяясь к фосфатидилинозитолгликану. Под действием фосфолипазы С может происходить отделение белков от внешней поверхности клетки.
Липиды могут быть аллостерическими активаторами мембранных ферментов. Например, β- гидроксибутиратдегидрогеназа, участвующая в окислении кетоновых тел (см. раздел 8), локализована на внутренней мембране митохондрий. Каталитическая активность фермента проявляется только в присутствии фосфатидилхолина.
Фермент протеинкиназа С катализирует реакции фосфорилирования белков по аминокислотным остаткам серина и треонина. В неактивной форме протеинкиназа С находится в цитозоле. Однако после стимуляции клетки (повышение в клетке концентрации кальция) фермент быстро активируется ионами кальция и оказывается связанным с мембраной. Функционально активная протеинкиназа С — комплекс, содержащий мономер фермента, молекулу диацилглицерола, один или более ионов Са2+ и четыре молекулы фосфатидилсерина.
Креатинкиназа, фермент катализирующий образование макроэргического соединения креатинфосфата (см. раздел 9). Для проявления его активности требуется специфическое взаимодействие с кардиолипином внутренней мембраны митохондрий.