Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ I. БІОМОЛЕКУЛИ ТА КЛІТИННІ СТРУКТУРИ
ГЛАВА 5. ЛІПІДИ. БІОМЕМБРАНИ
5.3. БІОЛОГІЧНІ МЕМБРАНИ
Біологічні мембрани (біомембрани) — клітинні структури, що відокремлюють клітину від навколишнього середовища та розділяють внутрішньоклітинний простір на певні компартменти (органели, субклітинні структури).
Протягом багатьох років основним науковим обґрунтуванням наявності на поверхні живих клітин спеціальних структурних утворень — мембран — був феномен обмеженої та вибіркової проникності клітини для хімічних сполук в іонній та молекулярній формах. У свою чергу, ця обмежена проникність зумовлює притаманну будь-якій клітині різницю концентрацій іонів усередині і в зовнішньому середовищі та наявність електричної різниці потенціалів між цитоплазмою і зовнішньоклітинною рідиною, особливо вираженої у збудливих нервових та м’язових клітинах.
Прямим доведенням наявності на поверхні клітини спеціального морфологічного утворення — плазматичної мембрани стали безпосередні електронно- мікроскопічні дослідження, які встановили також тришарову структуру всіх клітинних мембран, що відповідає внутрішньому ліпідному шару, вкритому ззовні та зсередини білковими молекулами. Електронномікроскопічне зображення плазматичної мембрани, що оточує еритроцит, подано на рис. 5.1.
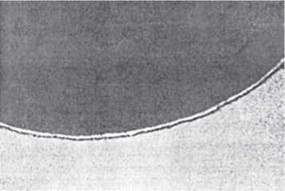
Рис. 5.1. Тришарова будова плазматичної мембрани еритроцита (за Е. Sim, 1982).
Функції біомембран:
а) відмежовування внутрішньоклітинного простору від навколишнього хімічного середовища за рахунок вибіркової проникності плазматичних мембран для іонів та молекул;
б) створення та підтримання на плазматичній мембрані іонних градієнтів та електричних потенціалів;
в) регуляція клітинних функцій біорегуляторними хімічними сигналами, що надходять від нервової та ендокринної систем;
г) поділ клітини на окремі компартменти, що характеризуються специфічними наборами ферментів, метаболітів та реакцій обміну речовин;
д) створення структурних, біофізичних умов для організації мембранозв’язаних мультиферментних комплексів (ферментних ансамблів), які реалізують життєво важливі клітинні функції (наприклад, електротранспортних ланцюгів у мембранах мітохондрій та ендоплазматичного ретикулуму), функціонування іонних каналів та насосів);
е) участь у процесах міжклітинної взаємодії як необхідного фактора регуляції клітинного росту, створення тканин (гістогенезу).
Мембранні структури тваринної клітини.
- плазматична мембрана;
- мембрани ендоплазматичного (саркоплазматичного) ретикулуму;
- мітохондріальні мембрани;
- ядерна мембрана;
- мембрани комплексу Гольджі;
- мембрани лізосом та фагосом;
- мембрани пероксисом (мікротілець).
Молекулярні компоненти біомембран
Головними хімічними компонентами біологічних мембран є білки, ліпіди та вуглеводи. Співвідношення між вказаними біохімічними компонентами значно відрізняється в окремих типах біомембран і залежить від їх функціональної та біохімічної спеціалізації (табл. 5.3).
Таблиця 5.3. Середній хімічний склад (%) деяких клітинних мембран
Білки |
Ліпіди |
Вуглеводи |
|
Плазматична мембрана еритроцитів людини |
49 |
43 |
8 |
Внутрішня мембрана мітохондрій печінки |
76 |
24 |
0 |
Мембрани ендоплазматичного ретикулума клітин печінки |
55 |
45 |
0 |
Мієлінові мембрани мозку людини |
18 |
79 |
3 |
Salmonella typhimurium |
|||
зовнішні мембрани |
44 |
20 |
36 |
внутрішні мембрани |
65 |
35 |
0 |
Як свідчать дані таблиці 5.3, вміст білків у клітинних мембранах становить в середньому 50-75 %, ліпідів — 25-45 %, вуглеводів — 0-10 %. Для зовнішніх клітинних (плазматичних) мембран притаманна наявність певної кількості вуглеводів, що входять до складу гліколіпідів та глікопротеїнів, для мієлінових мембран мозку - значна концентрація ліпідів. Внутрішні (субклітинні) мембрани мітохондрій, ендоплазматичного ретикулума містять відносно більшу кількість білків, що відображує наявність у цих мембранних структурах важливих мультиферментних комплексів.
А. Ліпіди мембран
Ліпідні компоненти біологічних мембран представлені переважно різними класами полярних ліпідів. фосфоліпідами (фосфатидилхоліном, фосфатидилетаноламіном, фосфатидилсерином, сфінгомієліном) — до 80-90% загального вмісту мембранних ліпідів; гліколіпідами (переважно глікосфінголіпідами). Зовнішня плазматична мембрана характеризується значним вмістом вільного холестеролу та його ефірів і наявністю гліколіпідів, які відсутні в інших мембранних структурах (табл. 5.4).
Таблиця 5.4. Ліпідний склад(%) субклітинних мембран печінки щурів (за А. Ленінджером, 1985)
Мембрана |
Фосфоліпіди |
Гліколіпіди |
Холестерин |
Ефіри холестеролу та інші мінорні ліпіди |
Плазматична |
57 |
6 |
15 |
22 |
Ядерна |
85 |
0 |
5 |
10 |
Мітохондріальна внутрішня |
92 |
0 |
0 |
8 |
Ендоплазматичного ретикулума |
85 |
0 |
5 |
10 |
Апарату Гольджі |
57 |
0 |
9 |
34 |
Особливістю структурної організації молекул ліпідів, що входять до складу біологічних мембран — фосфоліпідів та гліколіпідів, є наявність у них гідрофільної «голівки», що утворена залишком фосфату, етерифікованим полярними або зарядженими групами, та гідрофобних «хвостиків», які утворені ацилами насичених та ненасичених жирних кислот (С16, С18, С20 тощо) — рис. 5.2 та рис. 5.3.
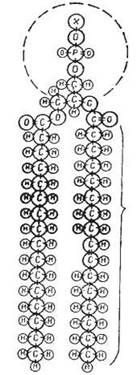
Рис. 5.2. Молекулярна будова фосфогліцеридів біологічних мембран.
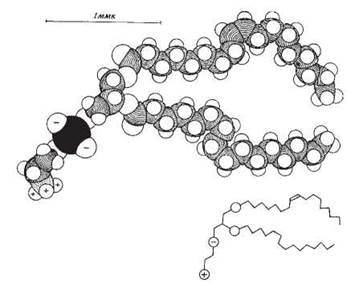
Рис. 5.3. Молекулярна модель фосфатидилетаноламіну
(відображено конформацію гідрофобних ацилів, яку вони набувають через наявність подвійних зв’язків).
На рис. 5.4 подано схематичні зображення молекулярних моделей основних мембранних ліпідів, у яких гідрофобний кінець молекул складають вуглеводневі радикали жирних кислот або вищого спирту сфінгозину, гідрофільний — іонізовані фосфати, ковалентно зв’язані з залишками холіну, етаноламіну, серину, гліцеролу, інозитолу, вуглеводу.
Молекула холестеролу входить до складу біомембран також завдяки наявності в її структурі гідрофобної частини (поліциклічний вуглеводень циклопентан-пергідрофенантрен) та гідрофільної — ОН-групи — рис. 5.5.
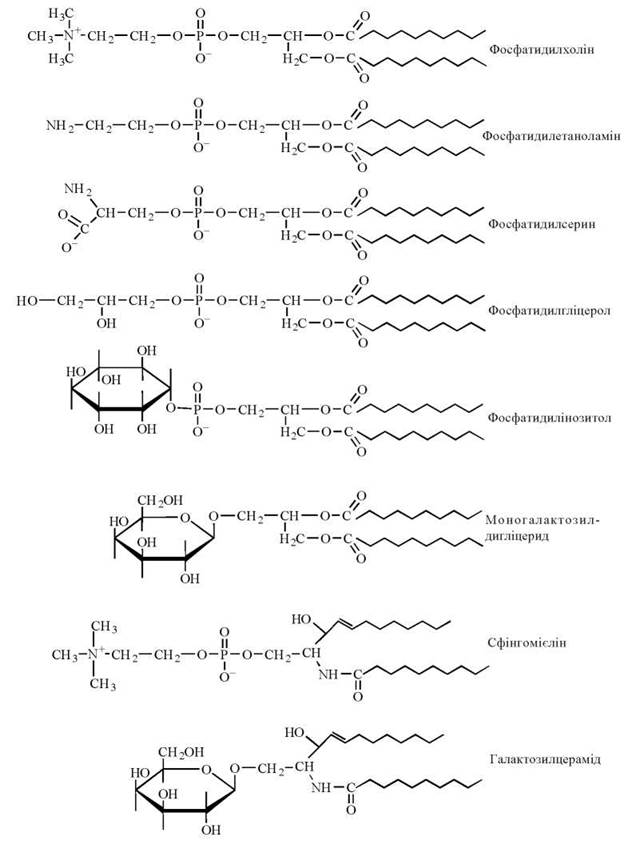
Рис. 5.4. Схематичні зображення молекулярних моделей ліпідів біомембран.
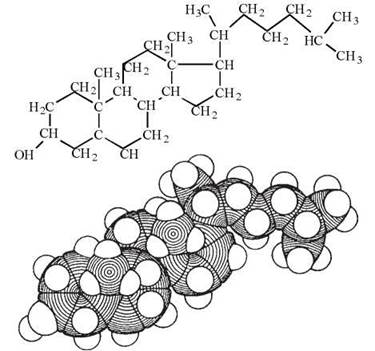
Рис. 5.5. Модель молекули холестеролу.
Б. Білки мембран
Білки біологічних мембран — це, переважно: ферменти; білки іонних каналів та інших систем мембранного транспорту; рецепторні білки, що зв’язують зовнішні ліганди та беруть участь у трансформації хімічного сигналу в біологічну реакцію клітини.
Певна кількість мембранних білків зв’язана з вуглеводами (глікозильована) у вигляді глікопротеїнів.
За характером розташування у мембрані білки поділяють на зовнішні (периферичні) та внутрішні. Зв’язок білків із різними мембранними структурами буде розглянуто нижче.
В. Вуглеводи мембран
Вуглеводи в складі біологічних мембран зв’язані з іншими хімічними компонентами мембрани у вигляді гліколіпідів та глікопротеїнів.
Гліколіпіди мембран є, головним чином, похідними сфінгозину (глікосфінголіпіди, або глікоцераміди).
Глікопротеїни мембран є молекулярними структурами, що утворюються за рахунок ковалентних зв’язків олігосахаридних ланцюгів з мембранними білками. Ці зв’язки формуються за участю гідроксильних груп серину або треоніну (О-глікозидні зв’язки) та амідної групи аспарагіну (N-глікозидний зв’язок).
Мономерними залишками у складі олігосахаридних ланцюгів мембранних гліколіпідів та глікопротеїнів є такі моносахариди та їх похідні: галактоза, глюкоза, маноза, галактозамін, глюкозамін, нейрамінова та сіалова кислота, фруктоза.
Гліколіпіди та глікопротеїни входять до складу, як правило, плазматичної мембрани клітини, контактуючи із зовнішньоклітинним оточенням та міжклітинним матриксом. Олігосахаридні залишки виконують функції лігандів для зовнішніх білків, тобто забезпечують процес розпізнавання та міжклітинної взаємодії, особливо важливі в реакціях клітинного імунітету. Аномальні зміни структури поверхневих гангліозидів у мембранах пухлинних клітин призводять до втрати характерного для росту нормальних клітинних пластів феномену «контактного гальмування», що супроводжується притаманним злоякісним пухлинам інфільтративним ростом.
Молекулярна організація біомембран
Наявність у мембранних ліпідах (гліцерофосфоліпідах, сфінгофосфоліпідах, гліколіпідах) полярних голівок та неполярних гідрофобних структур (вуглеводневих радикалів жирних кислот та сфінгозину) визначає їх амфіфільну (амфіпатичну) природу, тобто здатність до взаємодії як з гідрофільними (полярними), так і з гідрофобними (неполярними) молекулами.
Завдяки амфіфільній будові молекул, ліпіди, що беруть участь у побудові біомембран, здатні до утворення в полярних середовищах впорядкованих структур: міцел, моношарових та бішарових плівок (моношарів та бішарів) — рис. 5.6.

Рис. 5.6. Мембранні структури, що утворюються полярними ліпідами.
а) Міцели — молекулярні структури, які амфіфільні ліпіди утворюють у водному (полярному) оточенні. У міцелах вуглеводневі хвости ліпідів вкриті від контакту з водою та утворюють гідрофобну фазу, а гідрофільні голівки молекул розташовуються на поверхні. Міцелярні структури характерні для ліпопротеїнів крові та для ліпідних комплексів, що всмоктуються в кишковому тракті.
б) Мономолекулярні шари — плівки, які амфіфільні ліпіди утворюють на поверхні водних розчинів. У мономолекулярних шарах гідрофільні голівки молекул взаємодіють з водною фазою, а вуглеводневі хвости спрямовані до повітряної фази. Мономолекулярний шар, що утворює в легеневих альвеолах фосфогліцерид дипальмітоїлфосфатидилхолін, виконує функцію легеневого сурфактанту, який протидіє злипанню легеневих альвеол.
в) Бімолекулярні шари — молекулярні структури, в яких вуглеводневі хвости ліпідів спрямовані всередину, утворюючи неперервний вуглеводневий бішар, а гідрофільні (полярні) голівки направлені в бік водної фази, що оточує бімолекулярну плівку, яка утворилася; бімолекулярні шари є основою будови біологічних мембран.
Амфіфільний характер мембранних ліпідів є фізико-хімічною властивістю, що зумовлює їх здатність до утворення ліпідних бішарів, які складають основу молекулярної структури біологічних мембран.
Рідинно-мозаїчна модель будови біомембрани
Вперше припущення про те, що основу молекулярної організації біомембран складає подвійний ліпідний шар (бішар) було висунуте в 1925 р. Гортером та Гренделем (E. Gorter, F. Grendel). У 1935 р. Даніелі та Даусон (J.F. Danielli, H. Davson) запропонували модель, згідно з якою біологічні мембрани складаються з подвійного
шару ліпідів, який вкрито із зовнішнього та внутрішнього боку шарами білків — «бутербродна» модель. Модифікація моделі Даніелі-Даусона (модель Стейна-Даніелі, 1956) постулювала наявність в біомембранах полярних пор, придатних для трансмембранного переносу гідрофільних молекул — рис. 5.7.
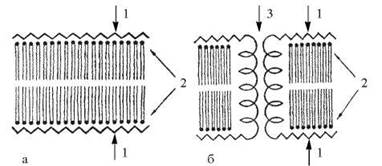
Рис. 5.7. Моделі біомембран за Даніелі-Даусоном (а) та Стейном-Даніелі (б): 1 — білкові шари; 2 — бішар ліпідів; 3 — мембранні пори.
Згідно з сучасною рідинно-мозаїчною моделлю Сінгера-Ніколсона (S.J. Singer, G.L. Nicolson), основу (безперервний матрикс) біологічної мембрани складає полярний ліпідний бішар, в який занурені окремі білкові молекули. За умов нормальних фізіологічних температур ліпіди біомембран знаходяться в рідинному стані, являючи собою «ліпідне озеро», в якому плавають, подібно до айсбергів, мембранні білки — рис. 5.8.
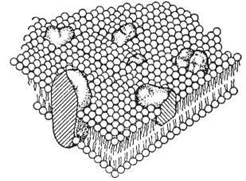
Рис. 5.8. Рідинно-мозаїчна модель будови біомембран.
За своєю локалізацією відносно інших компонентів біомембрани мембранні білки поділяються на такі типи (рис. 5.9):
а) поверхневі (периферичні) білки;
б) білки, що частково занурені у бішар;
в) внутрішні (інтегральні) білки.
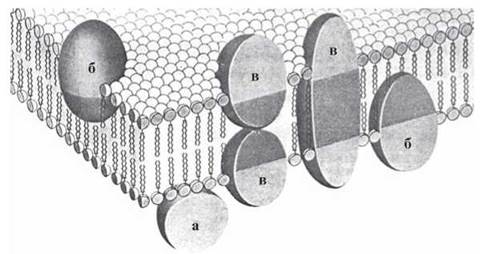
Рис. 5.9. Молекулярна організація біомембрани. Мембранні білки.
Біофізичні властивості мембран
1. Плинність та в’язкість ліпідної фази, що визначається співвідношенням між ненасиченими (рідкими) та насиченими (твердими) жирними кислотами в складі мембранних ліпідів та постійною рухомістю вуглеводневих хвостів ацилів та сфінгозину (подібно до «корзини із живими зміями») — рис. 5.10.
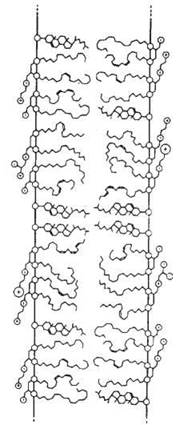
Рис. 5.10. Рухомість жирнокислотних залишків у молекулах мембранних ліпідів.
Холестерол, що входить до складу біомембран, виконує важливу функцію модифікатора фізико-хімічних властивостей ліпідного бішару, стабілізуючи його шляхом обмеження рухомості внутрішньомембранних компонентів, тобто зменшуючи плинність та збільшуючи в’язкість матриксу мембранних ліпідів.
2. Рухомість окремих молекулярних компонентів мембрани — ліпідів та білків.
Ліпіди біомембран мають певну впорядкованість, але разом з тим вони здатні до латеральної дифузії, тобто переміщення впродовж рідинної ліпідної фази (рідкісно-кристалічний стан мембранних ліпідів).
До латеральної дифузії здатні також молекули мембранних білків, що сприяє утворенню внутрішньо-мембранних білок-білкових ансамблів (кластерів). Важливим прикладом фізіологічного значення кластероутворення білків у площині біологічної мембрани є «шапко- утворення» (capping) мембранних рецепторів лімфоцитів при дії на клітину чужорідних лігандів.
3. Асиметрія мембранної структури.
Зовнішня та внутрішня поверхні будь-якої мембрани
суттєво відрізняються за своїми фізико-хімічними властивостями та за складом основних біохімічних компонентів, що пов’язано з різною функціональною спеціалізацією двох поверхонь мембран. Із зовнішньою поверхнею плазматичних мембран зв’язані рецептори для гормонів і інших фізіологічно активних речовин, із внутрішньою — деякі цитозольні ферменти та компоненти цитоскелета. Внутрішній моношар ліпідного бішару відрізняється від зовнішнього за складом фосфоліпідів.
Наприклад, зовнішня поверхня мембрани еритроциту містить олігосахаридні залишки гліколіпідів, які відіграють роль детермінант груп крові (система А, В, О); із зовнішньою поверхнею еритроцитарної мембрани зв’язаний фермент ацетилхолінестераза, із внутрішньою — білок спектрин — рис. 5.11.
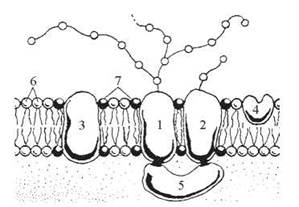
Рис. 5.11. Схема молекулярної організації плазматичної мембрани еритроцита (А.А. Заварзин, А.Д. Харазова, 1982):
1 — білок глікофорин; 2 — мембранний глікопротеїн; 3 — Na+, K+—АТФ-аза; 4 — ацетилхолінестераза; 5 — білок спектрин; 6 — ліпіди бішар; 7 — ліпіди, що контактують з мембранними білками.