Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ II. ЗАГАЛЬНІ ЗАКОНОМІРНОСТІ МЕТАБОЛІЗМУ
ГЛАВА 9. БІОЕНЕРГЕТИЧНІ ПРОЦЕСИ: ТРАНСПОРТ ЕЛЕКТРОНІВ; ОКИСНЕ ФОСФОРИЛЮВАННЯ В МІТОХОНДРІЯХ
9.4. ОКИСНЕ ФОСФОРИЛЮВАННЯ ТА АТФ-СИНТЕТАЗА МІТОХОНДРІЙ
Окисне фосфорилювання — процес, шляхом якого хімічна енергія, що вивільняється під час транспортування електронів упродовж дихального (електронотранспортного) ланцюга мітохондрій, уловлюється та використовується для синтезу аденозинтрифосфату (АТФ) з аденозиндифосфату (АДФ) та неорганічного фосфату (ФН).
Синтез АТФ з АДФ та неорганічного фосфату ФН(Pi — англ.) отримав назву спряження дихання (електронного транспорту в мітохондріях) та окисного фосфорилювання.
Вивільнення хімічної енергії в дихальному ланцюзі та ділянки утворення АТФ
Транспортування електронів у дихальному ланцюзі мітохондрій від первинних донорів відновлювальних еквівалентів (субстратів біологічного окислення) до молекулярного кисню супроводжується зниженням вільної енергії (ΔG), значення якого можна розрахувати за рівнянням:
![]()
де: п — кількість електронів, що транспортуються (дорівнює двом при утворенні молекули води);
F — число Фарадея [23,062 ккал/в · моль];
ΔE0' — різниця стандартних потенціалів електронодонорної та електроноакцепторної систем (при pH = 7,0).
Враховуючи дані таблиці 9.1, виконаємо розрахунок вивільнення хімічної енергії за умов перенесення двох електронів від піридинзалежних дегідрогеназ до кисню:
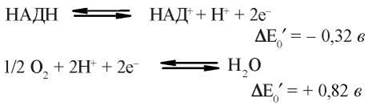
Сумування двох рівнянь дає:
![]()
що відповідає
![]()
Синтез однієї молекули АТФ з АДФ та ФН потребує витрат хімічної енергії, що дорівнюють + 7,3 ккал (+ 30,5 кДж). Очевидно, що енергії, яка вивільняється за умов транспорту електронів у дихальному ланцюзі мітохондрій, достатньо для синтезу декількох молекул АТФ. Безпосередніми біохімічними дослідженнями доведено, що за умов окислення субстратів через НАДН-коензим Q-редуктазу утворюється 3 молекули АТФ, при дії сукцинат-коензим Q-редуктази — 2 молекули АТФ.
Коефіцієнт окисного фосфорилювання — відношення кількості зв’язаного (етерифікованого) неорганічного фосфату (моль) до кількості поглинутого мітохондріями кисню (моль) (позначається як ФН (Р1)/О) — кількісно дорівнює числу молекул АТФ, що утворюються при перенесенні двох відновлювальних еквівалентів на один атом кисню, тобто АТФ/О — табл. 9.2.
Таблиця 9.2. Значення коефіцієнта окисного фосфорилювання при окисленні різних субстратів у мітохондріях
Субстрат |
Коефіцієнт ФН/О (АТФ/О) |
α-Кетоглутарат |
3 |
Ізоцитрат |
3 |
Малат |
3 |
L-Глутамат |
3 |
Сукцинат |
2 |
Ацил-КоА |
2 |
Пункти спряження транспорту електронів та окисного фосфорилювання
Утворення АТФ з АДФ та ФН може відбуватися тільки в певних ділянках електронотранспортного ланцюга мітохондрій, в яких величина хімічної енергії, що виділяється при транспортуванні пари електронів між двома редокс-системами (компонентами дихального ланцюга), достатня для синтезу 1 молекули АТФ (тобто > 7,3 ккал, або 30,5 кДж) — табл. 9.3.
Таблиця 9.3. Ділянки дихального ланцюга мітохондрій, де вивільнення хімічної енергії достатнє для синтезу молекули АТФ
Ділянка дихального ланцюга |
ΔE0' (вольт) |
ΔG |
|
(ккал) |
(кДж) |
||
Комплекс I (НАДН —► коензим Q) |
0,27 |
12,2 |
51,0 |
Комплекс III (цитохром b —► цитохром с) |
0,22 |
9,9 |
41,4 |
Комплекс IV (цитохром а3 —► О2) |
0,53 |
23,8 |
99,6 |
Зазначені ділянки електронотранспортного ланцюга називаються пунктами спряження дихання (електронного транспорту) з окисним фосфорилюванням — рисунок 9.3.
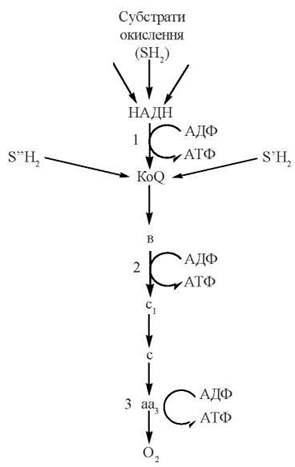
Рис. 9.3. Локалізація пунктів спряження (1,2,3) дихання та окисного фосфорилювання в електронотранспортному ланцюзі внутрішніх мембран мітохондрій.
Хеміоосмотична теорія окисного фосфорилювання
Молекулярні механізми генерації АТФ в ході біологічного окислення в мітоходріях пояснюються хеміоосмотичною теорією (за П. Мітчелом — P. Mitchell).

Рис. 9.4. Мітчел (Mitchell) Пітер Д. (народ. 1920 р.), англійський біохімік. Автор хеміоосмотичної теорії окисного та фотосинтетичного фосфорилювання. Нобелівська премія (1978).
Головний постулат хеміоосмотичної теорії — спряження електронного транспорту в мітохондріях із біохімічною системою синтезу АТФ здійснюється за рахунок електрохімічного потенціалу протонів (ΔμН+), що утворюється під час функціонування електронотранспортного ланцюга.
Хеміоосмотична теорія передбачає, що:
1. Функціонування дихального (електронотранспортного) ланцюга у внутрішніх (спрягаючих) мембранах мітохондрій супроводжується генерацією на цих мембранах електрохімічного градієнта протонів (Н+).
2. Окремі компоненти електронотранспортного ланцюга діють як протонні помпи, що спричиняють векторний (перпендикулярний площині мембрани) транспорт протонів, спрямований у напрямку «матрикс —► зовнішня поверхня мембрани».
3. Електрохімічний потенціал протонів на спрягаючих мембранах, який створюється завдяки дії протонних помп дихального ланцюга, є рушійною силою синтезу АТФ з АДФ та ФH.
4. Існує ферментна система, що використовує
енергію електрохімічного протонного потенціалу для синтезу АТФ за рахунок транслокації протонів через мітохондріальну мембрану в напрямку « зовнішня поверхня —► матрикс».
Ця ферментна система, яка замикає протонний цикл на спрягаючих мембранах мітохондрій — протонна АТФаза (Н+-АТФаза), або АТФ-синтетаза.
АТФ-синтетаза є білком з четвертинною структурою, що складається з декількох білкових субодиниць, які утворюють компоненти F0 та F1 (F0F1— АТФаза).
5. Будь-які фізичні, хімічні та біологічні фактори, що пошкоджують цілісність спрягаючих мембран мітохондрій та розсіюють енергію електрохімічного градієнта, порушують синтез АТФ, тобто виступають якроз’єднувачі транспорту електронів та окисного фосфорилювання.
Таким чином, згідно з хеміоосмотичною теорією, спряження між переносом електронів в дихальному ланцюзі та синтезом АТФ здійснюється за рахунок утворення при функціонуванні протонних помп градієнта концентрації Н+ між двома поверхнями мітохондріальної мембрани. АТФ-синтетаза, транспортуючи протони у зворотному напрямку (за електрохімічним градієнтом) призводить до вивільнення хімічної енергії, за рахунок якої утворюються макроергічні зв’язки АТФ.
Схему протонного циклу на спрягаючих мембранах мітохондрій як рушійної сили окисного фосфорилювання подано на рисунку 9.5.
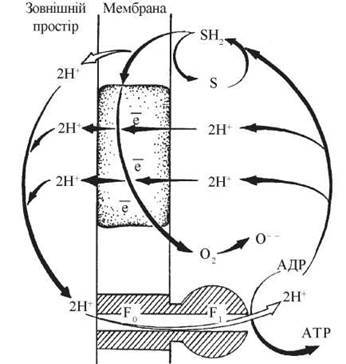
Рис. 9.5. Хеміоосмотичний механізм спряження транспорту електронів у дихальному ланцюзі з синтезом АТФ, АТФ-синтетазою за рахунок функціонування векторно спрямованих (перпендикулярно площині мембрани) протонних помп.
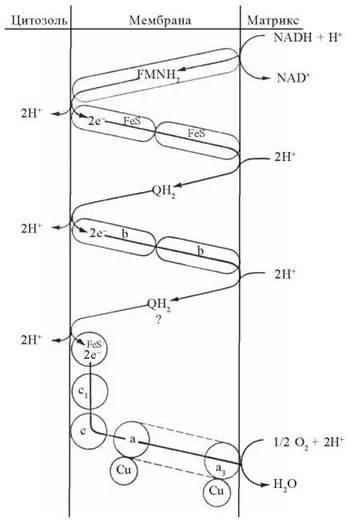
Рис. 9.6. Схема внутрішньомембранної організації електронотранспортних комплексів у вигляді «петель», що двічі перетинають мембрану.
Спроможність мітохондріальних переносників електронів до транслокації протонів через мембрану зумовлюється особливостями їх внутрішньомембранної топографії. Вважають, що дихальний ланцюг укладений у спрягаючій мембрані у вигляді трьох окислювально-відновлювальних «петель», що відповідають трьом комплексам переносу електронів — I, III та IV. Кожна пара відновлювальних еквівалентів (2Н+ + 2е-), проходячи через «петлю», транспортує два іони Н+ з матриксу в зовнішнє середовище. Зворотний перенос іонів Н+ через протонний канал АТФ-синтетази спрямовує електрохімічну енергію в бік синтезу АТФ:
![]()