Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ III. МЕТАБОЛІЗМ ОСНОВНИХ КЛАСІВ БІОМОЛЕКУЛ
ГЛАВА 14. МЕТАБОЛІЗМ ЛІПІДІВ. I. КАТАБОЛІЗМ ТРИАЦИЛГЛЩЕРОЛШ ТА ЖИРНИХ КИСЛОТ
14.2. КАТАБОЛІЗМ ТРИАЦИЛГЛІЦЕРОЛІВ
Триацилгліцероли (нейтральні жири, жири) потрапляють до організму людини як компоненти тваринної і рослинної їжі. Ліпіди цього класу розщеплюються в травному каналі до моногліцеридів, вільних жирних кислот та гліцеролу, які після всмоктування в кишечнику та складних біохімічних перетворень в ентероцитах, крові та печінці депонуються в жировій тканині. Накопичення резервів нейтральних жирів є найбільш ефективним механізмом акумулювання метаболічної енергії, оскільки завдяки високому ступеню відновлення жирнокислотних залишків, що входять до складу молекул триацилгліцеролів, при їх окисленні вивільняється значно більше хімічної енергії, ніж при катаболізмі вуглеводів та білків (глава 26).
Основне місце локалізації резервних тригліцеридів в організмі людини — адипоцити жирової тканини (ліпоцити), значна частина цитоплазми яких зайнята гігантською сферичною ліпідною краплею — рис. 14.1.
Рис. 14.1. Клітина жирової тканини (адипоцит).
У значно меншій кількості тригліцериди наявні в клітинах інших органів, зокрема гепатоцитів печінки, де вони перебувають у вигляді жирових краплин у цитоплазмі.
Жирова тканина, до 65 % маси якої припадає на нейтральні жири, є високоспеціалізованою тканиною, що акумулює значні кількості метаболічного палива, енергетична цінність якого суттєво перевищує енергетичні резерви вуглеводів і білків. Маса жирової тканини у дорослої людини середньої ваги дорівнює в середньому 10 кг, і загальний вміст тригліцеридів у ній достатній для забезпечення енергетичних потреб організму протягом, принаймні, 40 днів голодування.
Реакції катаболізму триацилгліцеролів
Ферментативний гідроліз (ліполіз) триацилгліцеролів в адипоцитах та інших клітинах, де накопичуються нейтральні жири, є фізіологічним механізмом, що має суттєве значення як резервне джерело енергії, особливо в умовах вичерпання вуглеводних резервів та при стресових ситуаціях. Процес розщеплення триацилгліцеролів із вивільненням жирних кислот, які виходять у кров, отримав назву мобілізації жирних кислот із жирової тканини.
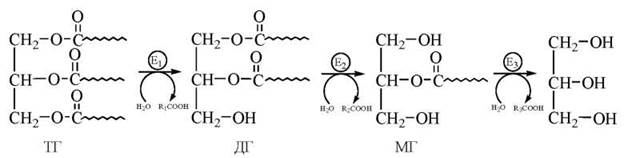
Внутрішньоклітинний ліполіз триацилгліцеролів (ТГ) здійснюється в декілька стадій, продуктами яких є діацилгліцероли (дигліцериди — ДГ), моноацилгліцероли (моногліцериди — МГ), гліцерол та вільні жирні кислоти:
Сумарне рівняння ліполізу:
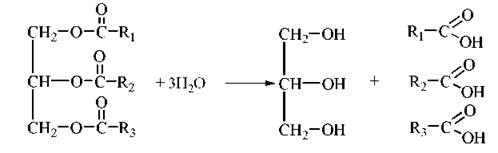
Вільні жирні кислоти (неестерифіковані жирні кислоти — НЕЖК) є субстратами окислення для клітин багатьох тканин, зокрема міокарда, хребцевої, гладенької мускулатури тощо, крім головного мозку. Після надходження в плазму крові нерозчинні у водній фазі плазми крові високомолекулярні жирні кислоти транспортуються у зв’язаній з сироватковим альбуміном молекулярній формі.
Молекулярні механізми регуляції ліполізу
Зазначений ступеневий процес ліполізу, що відбувається в адипоцитах жирової тканини, каталізується трьома ферментами — тригліцерид-, дигліцерид- та моногліцеридліпазою. Активність двох останніх ферментів (E2 та E3) в декілька десятків разів перевищує активність першого ферменту (E1). Звичайно загальна швидкість багатоступеневого метаболічного ланцюга контролюється активністю ферменту, що каталізує найбільш повільну (лімітуючу) стадію процесу. Тому такий фермент єрегуляторним, і, дісно, активність тканинної тригліцеридліпази (ТГ-ліпази) регулюється багатьма гормонами, зокрема адреналіном, глюкагоном, інсуліном, соматотропіном.
Молекулярної основою регуляції активності тригліцеридліпази адипоцитів є її ковалентна модифікація шляхом оберненого фосфорилювання — дефосфорилювання.
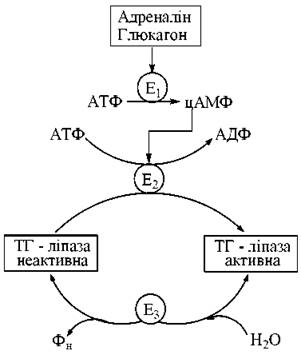
Рис. 14.2. Схема каскадної регуляції активності тригліцеридліпази адипоцитів.
Е1 — аденілатциклаза, Е2 — протеїнкіназа, Е3 — протеїнфосфатаза.
Фосфорильована форма ТГ-ліпази є каталітично активною, дефосфорильована — неактивною. Фосфорилювання відповідного білка здійснюється за рахунок АТФ при участі ферменту цАМФ-залежної протеїнкінази. У свою чергу, збільшення внутрішньоклітинної концентрації цАМФ є результатом взаємодії зі специфічними рецепторами плазматичних мембран адипоцитів адреналіну або глюкагону, що призводить до активації мембранозв’язаної аденілатциклази. Дефосфорилювання каталітично активної ТГ-ліпази фосфатазою призводить до утворення неактивної молекулярної форми ферменту.
У цілому процес контролю активності ТГ-ліпази в жировій тканині є прикладом залежного від цАМФ каскадного механізму гормональної регуляції біохімічних та фізіологічних функцій клітини, що був раніше розглянутий (глава j3) на прикладі фосфоролізу глікогену в печінці та м’язах.
Нейрогуморальна регуляція ліполізу
Регуляція метаболізму ліпідів здійснюється за рахунок впливу фізіологічних та біохімічних ефектів нейроендокринної системи відносно швидкістьлімітуючих реакцій обміну даного класу біомолекул. Певні гормони та нейромедіатори впливають на каталітичну активність і внутрішньоклітинну концентрацію ключових ферментів розщеплення й біосинтезу ліпідів. Найбільш вивченою є нейрогуморальна регуляція ліполізу — внутрішньоклітинного гідролізу резервних триацилгліцеролів.
Адреналін, норадреналін, глюкагон
Адреналін та норадреналін — катехоламіни, що активують ліполіз у жировій тканині за рахунок стимуляції цАМФ-залежного каскадного механізму регуляції активності ТГ-ліпази адипоцитів. Ліполітична дія цих гормонів реалізується за умов фізіологічних (фізичне напруження, зниження температури навколишнього середовища) та психологічних (страх, тривога) стресів, що супроводжуються вивільненням з мозкового шару наднирникових залоз адреналіну, а також стимуляцією симпатичної нервової системи та вивільненням у синапсах нейронів норадреналіну, що взаємодіють із адренергічними рецепторами мембран адипоцитів.
Глюкагон — панкреатичний гормон, що стимулює ліполітичну систему в жировій тканині за механізмом, подібним до дії катехоламінів, тобто за рахунок підвищення в адипоцитах вмісту цАМФ, пов’язаного з активацією аденілатциклази. Дія глюкагону проявляється в умовах зниження концентрації глюкози в крові через зменшення її надходження з кишечника або посиленого використання в тканинах.
У цілому за рахунок розглянутих біохімічних механізмів метаболічні ефекти катехоламінів та глюкагону призводять до швидкої стимуляції глікогенолізу в печінці і м’язах та ліполізу в жировій тканині, що забезпечує підвищені енергетичні потреби організму за умов стресу або голодування.
Інсулін
На відміну від зазначених гуморальних факторів, що активують ТГ-ліпазу адипоцитів, спричиняючи мобілізацію НЕЖК із жирової тканини, гормон інсулін гальмує процес ліполізу та вивільнення жирних кислот. Інгібіруюча дія інсуліну відносно ліполізу в адипоцитах реалізується за рахунок двох біохімічних механізмів:
а) зменшення концентрації цАМФ, що може бути пов’язаним з активацією фосфодіестерази цАМФ;
б) збільшення проникності мембран адипоцитів до глюкози, результатом чого є активація в жировій тканині гліколізу і, відповідно, накопичення гліколітичних метаболітів діоксіацетонфосфату та 3-фосфогліцеринальдегіду. Ці метаболіти, в свою чергу, є попередниками гліцерол-3-фосфату, що необхідний для реетерифікації жирних кислот при біосинтезі триацилгліцеролів. Таким чином, стимульоване інсуліном підвищене надходження в адипоцити глюкози переключає метаболізм жирних кислот на використання їх здебільшого в синтетичних реакціях і зменшує їх вихід у кров.
Розглянуті біохімічні особливості дії інсуліну пояснюють певні зміни вуглеводного та ліпідного обмінів, що спостерігаються при голодуванні та цукровому діабеті. Зазначені стани, для яких властиве зниження концентрації інсуліну в крові, характеризуються також зниженням надходження в адипоцити глюкози, що, зменшуючи глюкозозалежне гальмування мобілізації жирних кислот (див. вище), сприяє їх виходу в плазму крові та використанню іншими тканинами як енергетичного джерела. Введення глюкози та інсуліну хворим на цукровий діабет або експериментальним тваринам, що голодують, спричиняє інгібірування ліполізу в адипоцитах і протидіє надмірному збільшенню НЕЖК у плазмі крові.
Соматотропін — гормон передньої частки гіпофіза, який також стимулює ліполіз у жировій тканині за умов голодування, але його ліполітична дія суттєво відрізняється від дії катехоламінів та глюкагону. Соматотропін спричиняє підвищення процесів ліполізу за рахунок підвищення синтезу відповідних ферментних білків. Метаболічні ефекти соматотропіну розвиваються повільно, що свідчить про його значення в поступовій адаптації до голодування.
Ліполіз в інших тканинах (м’язах, печінці) регулюється за подібними нейрогуморальними механізмами.