Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ III. МЕТАБОЛІЗМ ОСНОВНИХ КЛАСІВ БІОМОЛЕКУЛ
ГЛАВА 14. МЕТАБОЛІЗМ ЛІПІДІВ. I. КАТАБОЛІЗМ ТРИАЦИЛГЛЩЕРОЛШ ТА ЖИРНИХ КИСЛОТ
14.3. ОКИСЛЕННЯ ЖИРНИХ КИСЛОТ ТА ГЛІЦЕРОЛУ
Продуктами ліполізу триацилгліцеролів та інших гліцеридів є жирні кислоти і гліцерол, що здатні до окислення з генерацією значної кількості АТФ.
Окислення жирних кислот
Окислення жирних кислот відбувається в матриксі мітохондрій у результаті циклічного процесу, який включає в себе послідовне відщеплення від довголанцюгових молекул насичених жирних кислот, що складаються з парної кількості вуглецевих атомів (пальмітинової — С16, стеаринової — С18 тощо), двовуглецевих фрагментів — циклу β-окислення.
Як і в разі метаболізму глюкози, передумовою входження жирної кислоти на шлях окислення є її ферментативна активація, тобто перетворення в активне похідне в результаті реакції, що потребує використання молекули АТФ.
Активація жирних кислот відбувається в цитоплазмі за участю специфічних ферментів ацил-КоА-синтетаз (тіокіназ), що утворюють КоА-похідні жирних кислот:
![]()
Ферментативні реакції β-окислення жирних кислот
1. Дегідрування КоА -похідних жирних кислот за участю ФАД-залежного ферменту ацил-КоА-дегідрогенази.
У результаті реакції утворюється трансненасичене (в положеннях 2,3, або α, β) КоА-похідне жирної кислоти:
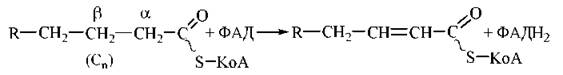
2. Гідратація ненасиченого КоА-ацилу ферментом еноїл-КоА-гідратазою з утворенням спиртового похідного ацил-КоА — 3-оксіацилу-КоА (β-гідроксіацилу-КоА):
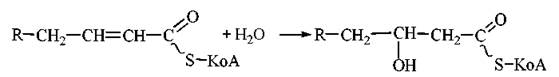
3. Дегідрування оксипохідного ацил-КоА НАД-залежним ферментом 3-оксіацил-КоА-дегідрогеназою. Продукт реакції — 3-кетоацил-КоА (β-кетоацил-КоА):
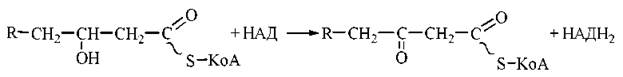
4. Тіолітичне розщеплення 3-кетоацил-КоА за рахунок взаємодії з молекулою КоА при участі ферменту β-кетоацил-КоА-тіолази. В результаті реакції утворюється молекула КоА-похідного жирної кислоти, скороченого на два вуглецеві атоми, та ацетил-КоА:
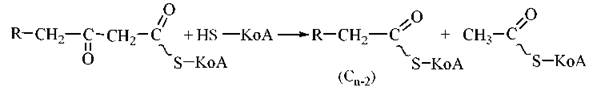
У результаті одного циклу β-окислення з молекули жирної кислоти вивільняється одна молекула ацетил-КоА і, відповідно, вихідна молекула ацил-КоА скорочується на два вуглецевих атоми. Легко зрозуміти, що для повного розщеплення до ацетил-КоА будь-якої молекули жирної кислоти з парною кількістю вуглецевих атомів (n) потрібно (n/2 - 1) циклів β-окислення.
Виходячи із зазначеного, сумарне рівняння β-окислення поширеної в природних триацилгліцеролах пальмітинової кислоти має вигляд:
![]()
Загальну схему циклу β-окислення насичених жирних кислот подано на рис. 14.3.
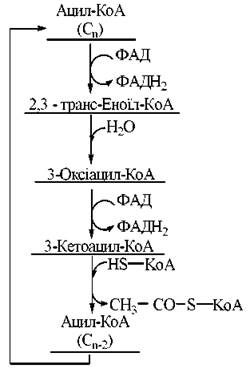
Рис. 14.3. Метаболічна карта процесу β-окислення жирних кислот.
Роль карнітину в окисленні жирних кислот
Ферменти β-окислення жирних кислот локалізовані всередині мітохондрій, але внутрішня мітохондріальна мембрана непроникна для довголанцюгових ацильних похідних КоА. Тому на внутрішній мітохондріальній мембрані функціонує спеціальна транспортна система, що включає аміноспирт карнітин, який бере участь у перенесенні молекул ацил-КоА до мітохондріального матриксу.
Транспортна функція карнітину реалізується за човниковим принципом (рис. 14.4.):
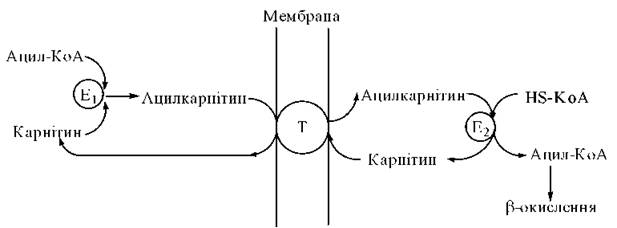
Рис. 14.4. Участь карнітину в перенесенні довголанцюгових жирних кислот через внутрішню мембрану мітохондрій. Е1 — карнітин-ацилтрансфераза I; Е2 — карнітин-ацилтрансфераза II; T — транслоказа.
а) на зовнішній поверхні внутрішньої мітохондріальної мембрани за участю ферменту карнітин-ацилтрансферази I відбувається утворення ефіру ацилкарнітину:
![]()
б) транспортний білок карнітин-ацилкарнітин-транслоказа переносить ацилкарнітин через мембрану мітоходрій;
в) на внутрішній поверхні мембрани фермент карнітин-ацилтрансфераза II розщеплює ацилкарнітин у наступній реакції:
![]()
Ацил-S-KoA вступає на шлях β-окислення, а вільний карнітин виходить з мітохондрій і бере участь у транспортуванні нової молекули жирної кислоти.
Енергетика β-окислення жирних кислот
1. У кожному циклі β-окислення вивільняється одна молекула ацетил-КоА, окислення якої в циклі трикарбонових кислот супроводжується утворенням 12 молекул АТФ (глава 10). β-Окислення пальмітату (див. вище) призводить до утворення 8 молекул ацетил-КоА, повне окислення яких до СО2 та Н2О дасть 96 (12x8) молекул АТФ.
2. У кожному циклі в-окислення утворюються дві молекули відновлених коферментів — ФАДН2 та НАДН, які можуть віддавати свої відновлювальні еквіваленти ланцюга електронного транспорту в мітохондріях, сприяючи генерації в результаті окисного фосфорилювання 2 (ФАДН2) та 3(НАДН), тобто сумарно 5 молекул АТФ. У разі повного окислення пальмітату в 7 циклах β-окислення за рахунок даного механізму утвориться 35 (5x7) молекул АТФ.
Враховуючи витрату 1 молекули АТФ на етапі активації жирної кислоти, загальна кількість молекул АТФ, що може синтезуватися в умовах повного окислення до діоксиду вуглецю та води молекули пальмітату, дорівнює 130 (96+35-1). Виходячи з цього, можна подати сумарне рівняння окислення пальмітинової кислоти в мітохондріях:
![]()
Окислення гліцеролу
Гліцерол, що утворюється при розщепленні триацилгліцеролів або гліцерофосфоліпідів, може вступати на шлях катаболізму (окислення) або знову використовуватися для біосинтезу різних класів гліцеридів (глава 15).
1. Включенню гліцеролу до метаболічних перетворень передує його активація, яка полягає в його трансформації за участю АТФ до гліцерол-3-фосфату (α-гліцерофосфату) при дії ферменту гліцеролфосфокінази.
2. α-Гліцерофосфат здатний до окислення мітохондріальним ферментом α-гліцерофосфатдегідрогеназою з утворенням гліцеральдегід-3-фосфату (Г-3-Ф).
Гліцеральдегід-3-фосфат є одним з центральних метаболітів гліколітичного окислення глюкози. Подальше перетворення Г-3-Ф, утворенного при окисленні гліцеролу, співпадає з катаболізмом гліколітичного Г-3-Ф (глава 11):

![]()