Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ III. МЕТАБОЛІЗМ ОСНОВНИХ КЛАСІВ БІОМОЛЕКУЛ
ГЛАВА 14. МЕТАБОЛІЗМ ЛІПІДІВ. I. КАТАБОЛІЗМ ТРИАЦИЛГЛЩЕРОЛШ ТА ЖИРНИХ КИСЛОТ
14.4. БІОСИНТЕЗ ТА КАТАБОЛІЗМ КЕТОНОВИХ ТІЛ
В умовах нормального метаболізму здорового організму основним шляхом використання ацетил-КоА, що утворюється при β-окисленні жирних кислот, є цикл трикарбонових кислот. В умовах переключення метаболізму на біосинтетичні шляхи цитоплазматичний ацетил-КоА знову використовується для формування жирних кислот, тобто утворення резервів ліпідів (глава 15).
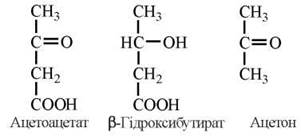
Разом з тим, у печінці існує фізіологічно важливий шлях утилізації ацетил-КоА, що призводить до утворення молекул альтернативного метаболічного палива, які використовуються в інших тканинах — так званих кетонових (ацетонових) тілах. До кетонових тіл належать ацетоацетат, β-гідроксибутират та ацетон.
Ферментні реакції утворення кетонових тіл
Утворення кетонових тіл відбувається в цитозолі (початкові етапи) та мітохондріях гепатоцитів за рахунок таких реакцій:
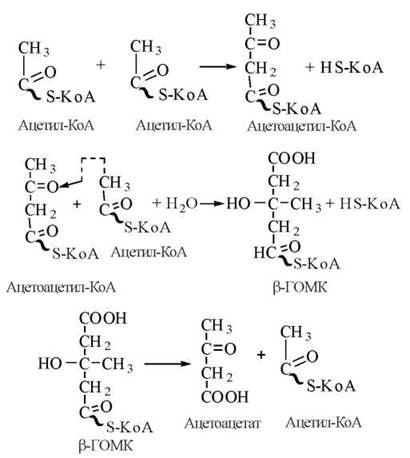
1. Конденсація двох молекул ацетил-КоА з утворенням ацетоацетил-КоА. Реакція каталізується цитозольним ферментом тіолазою.
2. Взаємодія ацетоацетил-КоА з новою молекулою ацетил-КоА з утворенням β-гідрокси-β-метилглутарил-КоА (β-ГОМК). При утворенні кетонових тіл реакція відбувається в мітохондріях і каталізується ферментом β-ГОМК-синтетазою.
3. Розщеплення β-гідрокси-β-метилглутарил-КоА мітохондріальним ферментом β-ГОМК-ліазою з утворенням ацетоацетату та ацетил-КоА.
Необхідно зауважити, що утворення β-ГОМК із ацетоацетил-КоА є процесом, який може здійснюватися в цитозолі гепатоцитів; у цьому випадку β-ГОМК підлягає біохімічним перетворенням, які становлять метаболічний шлях біосинтезу холестерину (глава 15), що відбувається в клітинах печінки, кишечника та шкіри. Однак у гепатоцитах більша частина ацетил-КоА, що утворюється при окисленні вуглеводів, жирних кислот, амінокислот і не використовується в цитратному циклі, вступає на шлях утворення кетонових тіл.
β-Гідроксибутират утворюється з ацетоацетату шляхом відновлення НАД-залежною β-гідроксибутиратдегідрогеназою:
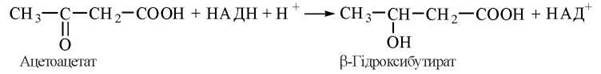
Реакція перебігає в напрямку утворення β-гідроксибутирату за умов високого співвідношення в гепатоцитах НАДН/НАД+, яке буває в умовах голодування.
Ацетон утворюється в незначній кількості з ацетоацетату, що міститься в циркулюючій крові, за рахунок його неферментативного декарбоксилювання або дії ферменту ацетоацетатдекарбоксилази. Ацетон видаляється з організму легенями; значне збільшення вмісту ацетону у видихуваному повітрі спостерігається в умовах декомпенсованого цукрового діабету.
Реакції утилізації кетонових тіл
Після утворення в гепатоцитах кетонові тіла (переважно ацетоацетат) виходять у кров і транспортуються в периферичні тканини, де вони виступають як важливі субстрати біологічного окислення.
Використанню ацетоацетату як субстрату метаболічного палива передує його активація з утворенням ацетоацетил-КоА. Існує два ферментативні механізми генерації ацетоацетил-КоА в гепатоцитах:
а) взаємодія ацетоацетату з сукциніл-КоА:
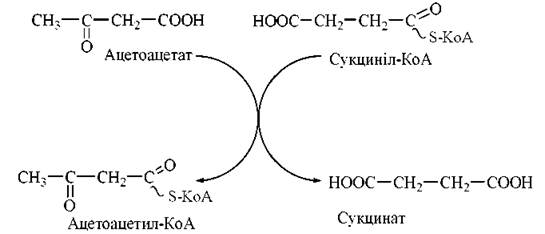
б) активація ацетоацетату HS-KoA за участю АТФ:
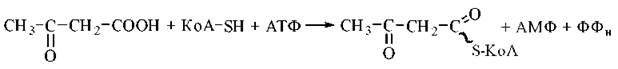
Ацетоацетил-КоА, що утворився за одним із зазначених механізмів, підлягає тіолітичному розщепленню за участю тіолази з утворенням двох молекул ацетил-КоА, які окислюються в циклі трикарбонових кислот:
![]()
Ацетоацетат є важливим джерелом енергетичного палива для міокарда, скелетних м’язів та коркового шару нирок, в клітинах яких катаболізм цієї сполуки перевищує утилізацію глюкози. В умовах голодування ацетоацетат стає переважаючим субстратом окислення також для головного мозку, який в нормальних умовах використовує для енергетичних потреб виключно глюкозу.
Метаболізм кетонових тіл в умовах патології
У нормі концентрація кетонових тіл у крові та більшості тканин незначна (в середньому 10-20 мг/л). Проте, за умов голодування та цукрового діабету створюються метаболічні умови, за яких кількість кетонових тіл у тканинах різко підвищується за рахунок значної активації їх синтезу. При цьому значно зростають як концентрація кетонових тіл у крові (кетонемія), так і їх виділення з сечею (кетонурія).
Біохімічною основою зростання вмісту кетонових тіл в умовах патології є зменшення ступеня утилізації ацетил-КоА в циклі трикарбонових кислот внаслідок порушень вуглеводного обміну.
Справа в тому, що входження ацетил-КоА в ЦТК залежить від наявності в клітині достатньої кількості оксалоацетату, необхідного для утворення цитрату. В свою чергу, утворення оксалоацетату, необхідного для нормального функціонування трикарбонового циклу, залежить від кількості пірувату (глава 10, розділ 10.4), основним постачальником якого є гліколітичне розщеплення глюкози. В умовах зменшеного надходження в клітину глюкози (голодування, цукровий діабет) оксалоацетат спрямовується на шлях глюконеогенезу і стає недосяжним для взаємодії з ацетил-КоА в цитратсинтазній реакції. У зазначених метаболічних умовах ацетил-КоА в значній мірі використовується для утворення кетонових тіл — ацетоацетату та β-гідроксибутирату. Сприяє накопиченню в клітинах ацетил-КоА також його підвищене утворення при β-окисленні жирних кислот за рахунок стимуляції в умовах глюкозного голодування ліполізу в жировій тканині. Ці біохімічні закономірності пояснюють давній вислів «Жири згоряють у полум ’ї вуглеводів».
Введення в організм глюкози (при голодуванні), або глюкози з інсуліном (при цукровому діабеті) підвищує внутрішньоклітинний рівень моносахариду і нормалізує гліколіз, що призводить до активації утилізації ацетил-КоА в циклі трикарбонових кислот і зменшення утворення кетонових тіл. Проте, в умовах відсутності необхідної терапії концентрація ацетоацетату, β-гідроксибутирату та ацетону в організмі хворих на цукровий діабет може зростати в десятки разів, супроводжуючись порушенням кислотно-лужного балансу і розвитком метаболічного кетоацидозу, який є небезпечним для нормального функціонування клітин головного мозку.