Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ IV. МОЛЕКУЛЯРНІ МЕХАНІЗМИ СПАДКОВОСТІ ТА РЕАЛІЗАЦІЇ ГЕНЕТИЧНОЇ ІНФОРМАЦІЇ
ГЛАВА 22. РЕГУЛЯЦІЯ ЕКСПРЕСІЇ ГЕНІВ. ГЕНЕТИЧНІ РЕКОМБІНАЦІЇ
22.3. МОЛЕКУЛЯРНІ МЕХАНІЗМИ МУТАЦІЙ. РЕПАРАЦІЯ ДНК
Мутації — зміни спадкових властивостей внаслідок кількісних та якісних змін у генотипі організму. В процесі реплікації ДНК мутації передаються від клітини до клітини і від покоління до покоління.
Разом з генетичними рекомбінаціями (див. вище), мутації складають основу спадкової мінливості живих організмів. Мутації можуть бути спричиненими певними природними — спонтанні мутації або штучними факторами — індуковані мутації.
За характером змін у структурі генетичного апарату організму мутації поділяють на:
(1) геномні мутації — такі, що полягають у змінах кількості повного набору хромосом або окремих хромосом у диплоїдному наборі; такі мутації спричиняють найбільш поширені та важкі форми хромосомних хвороб людини;
(2) хромосомні мутації — мутації, що пов’язані із структурними змінами певних хромосом (хромосомні аберації) внаслідок переміщення, втрати або дуплікації окремих фрагментів хромосомної ДНК. Розрізняють такі типи хромосомних мутацій:
транспозиції — перенесення фрагмента ДНК в іншу ділянку тієї ж хромосоми;
транслокації — перенесення ділянки однієї хромосоми на іншу, негомологічну їй, хромосому;
інверсії — зміна в певній ділянці хромосоми послідовності генів (азотистих основ) на іншу, зворотну послідовність;
делеції — випадіння певних ділянок хромосоми (фрагментів ДНК);
дуплікації — подвоєння певних ділянок хромосом.
(3) генні (точкові) мутації — зміни в структурі геному, що полягають в порушеннях послідовності азотистих основ (нуклеотидів), які складають первинну структуру ДНК. Генні мутації поділяють на такі типи:
а) заміни нуклеотидів — найбільш поширені генні мутації, до яких належать такі субтипи, як:
транзиції — заміна однієї пуринової основи на пуринову або піримідинової на піримідинову;
трансверзії — заміна одного типу азотистих основ на інший, тобто пурину на піримідин або навпаки;
б) випадіння (делеції) в ланцюгу ДНК однієї або декількох азотистих основ (і відповідних нуклеотидів);
в) вставки (вбудовування) в ланцюг ДНК додаткових азотистих основ (однієї або більшої кількості).
Генні мутації, якщо вони не репаровані спеціальними ферментними системами клітини, призводять до припинення синтезу білка, що кодується відповідним геном, або до утворення білка із зміненою, «неправильною» первинною структурою.
Агенти, що спричиняють мутації (мутагени)
Мутації (найчастіше генні мутації) виникають внаслідок пошкоджень, спричинених несприятливою дією на геном хімічних, фізичних та біологічних факторів навколишнього середовища, або похибок у функціонуванні ДНК-полімераз на етапі реплікації ДНК.
Найбільш поширеними мутагенами є:
(1) аналоги азотистих основ — сполуки, що заміщують нормальні азотисті основи в полідезоксирибонуклеотидному ланцюгу. Найбільш поширеними речовинами цього класу є 5-бромурацил та 2-амінопурин;
(2) хімічні мутагени — сполуки, що призводять до змін ковалентної структури нормальних азотистих основ; до найпоширеніших хімічних мутагенів належать:
а) дезамінуючі агенти — азотиста кислота (HNO2) та речовини, що в процесі метаболізму можуть перетворюватися на нітрити, зокрема органічні сполуки — нітрозаміни. Під дією азотистої кислоти відбувається дезамінування цитозину (з утворенням урацилу), аденіну і гуаніну (з утворенням гіпоксантину та ксантину, відповідно). Заміна одного нуклеотиду в ланцюгу ДНК супроводжується зміною змісту певного кодону (місенс-мутація) та синтезу білка із зміненою амінокислотною послідовністю. Вважають, що за рахунок мутації такого типу виникли аномальні форми гемоглобінів із зміненою первинною структурою в β-ланцюгах;
б) алкілюючі агенти — сполуки, що призводять до метилювання (в загальному випадку — алкілювання) звичайних азотистих основ. До алкілюючих агентів належать: алкілсульфонати (диметилсульфонат, етилметансульфонат тощо), азотисті та сірчанисті біс-(β-хлоретил)аміни (іприти), алкілнітрозаміни тощо; багато з цих сполук мають протипухлинну (антибластомну) активність і застосовуються в клінічній та експериментальній онкології;
(3) ультрафіолетове (УФ-) та іонізуюче опромінення — фізичні фактори, висока мутагенна активність яких пояснюється вільно-радикальною деструкцією азотистих основ ДНК з утворенням їх аналогів із зміненою хімічною будовою.
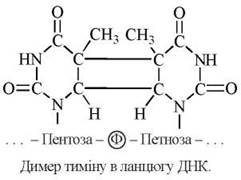
Поширеною мутацією, що спостерігається при дії УФ-випромінення є утворення ковалентних зв’язків між сусідніми (розташованими в одному ланцюгу) залишками тиміну. Такі тимінові димери протидіють нормальному просуванню ДНК-полімераз в ході реплікації і синтез ДНК припиняється.
Механізми репарації ДНК
Усі живі організми на Землі підлягають постійній дії фізичних мутагенних факторів, зокрема опроміненню УФ -променями (з довжиною хвилі 200-400 нм), що складають значну частину сонячного спектра, та іонізуючою радіацією, джерелом якої є космічне випромінення та радіоактивні ізотопи радію, плутонію, вуглецю тощо, що містяться в неорганічних об’єктах навколишнього середовища. За оцінками фахівців, вплив УФ - та іонізуючого опромінення є відповідальним за приблизно 10 % усіх пошкоджень ДНК, що спричиняються небіологічними факторами. Наступним важливим компонентом мутагенного впливу навколишнього середовища на геном живих організмів є дія численних хімічних сполук, зокрема речовин чужорідного походження — ксенобіотиків, які постійно впливають на ядерну генетичну ДНК. Як і в разі дії розглянутих фізичних чинників, механізм ушкоджуючої дії хімічних мутагенів значною мірою залежить від утворення в клітині вільних радикалів кисню та води, що спричиняють зміни в ковалентній структурі азотистих основ ДНК (Ю.І. Губский, 1993).
Вплив на геном живих організмів розглянутих фізичних та хімічних чинників супроводжується нестабільністю ДНК, що зазнає постійних точкових мутацій, найчастішими з яких є відщеплення пуринових основ (депуринізація ДНК), дезамінування цитозину та депіримідинізація. У зв’язку із зазначеним, в еволюції живих організмів виникли спеціальні молекулярні механізми, що протидіють постійним ушкодженням молекул ДНК та репарують зміни в молекулярній будові ДНК, що вже відбулися.
1. Репарація пошкоджень, спричинених УФ-опроміненням.
Пошкодження ДНК, спричинені УФ -променями, найбільш часто спостерігаються в бактеріальних клітинах та в незахищеній від сонячного опромінення шкірі людини. Відновлення нормальної структури ДНК, порушеної утворенням димерів тиміну, реалізується шляхом дії механізмів, позначених на рис. 22.14 (етапи I-IV):
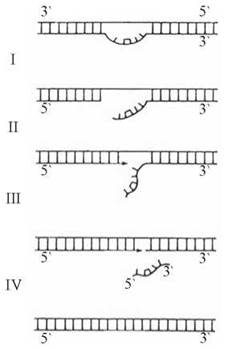
Рис. 22.14. Схема репарації фрагмента ДНК з тиміновим димером.
I. Розщеплення («розрізання») ланцюга ДНК «зліва» (в напрямку →5') від димера і відведення вбік вільного кінця, що містить тиміновий димер; реакція каталізується особливим ферментом — УФ-специфічною ендонуклеазою.
II. Формування полідезоксирибонуклеотидної «латки» на фрагмент ділянки ДНК, що містить димер; реакція каталізується ДНК-полімеразою (у прокаріотів — ДНК-полімеразою I) і полягає в приєднанні мононуклеотидів до вільного 3'-кінця «розрізаного» ланцюга ДНК в напрямку 5'→3'.
III. Відщеплення пошкодженої (такої, що містить тиміновий димер) ділянки ДНК (у прокаріотів — за рахунок 5'→3' єкзонуклеазної активності ДНК-полімерази I).
IV. «Зшивання» 3'-кінця новосинтезованої «латки» з 5'-кінцем розрізаного основного ланцюга ДНК.
Порушення ферментативного процесу репарації УФ-індукованих пошкоджень ДНК призводить у людини до важкого спадкового захворювання — пігментної ксеродерми (рис. 22.15).
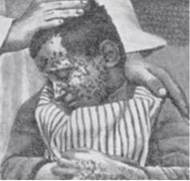
Рис. 22.15. Пігментна ксеродерма — сублетальне спадкове захворювання шкіри (A. Muntzing, 1967).
Пігментна ксеродерма успадковується як автосомальна рецесивна хвороба; при цій патології шкіра пацієнтів є надзвичайно чутливою до пошкоджуючої дії сонячного світла, яке може спричиняти розвиток раку шкіри. Найбільш поширена форма пігментної ксеродерми зумовлена спадковим порушенням синтезу УФ-специфічної ендонуклеази, що призводить до порушення всього механізму репарації ДНК.
2. Репарація дезамінування цитозину.
Дезамінування цитозину (2-окси-4-амінопіримідину) призводить до утворення урацилу (2,4-діоксипіримідину) — реакція, що спонтанно проходить у водних розчинах цитозину. В результаті цього процесу (заміни C на U) в комплементарному ланцюгу ДНК при реплікації також відбувається заміна відповідної пуринової основи (замість гуаніну включається аденін). У кінцевому підсумку дочірня дволанцюгова ДНК в тому положенні, де повинна була б бути пара G-C буде містити пару A-T, тобто відбудеться мутація типу заміни. Репарація такої мутації включає в себе: 1967).
а) видалення з ланцюга ДНК «неправильної» основи за рахунок відщеплення урацилу урацил-ДНК-глікозидазою. Фермент гідролітичним шляхом розриває N-глікозидний зв’язок між азотистою основою (урацилом) та дезоксирибозою, утворюючи пентозо-фосфатний ланцюг без азотистої основи. За аналогічним механізмом діють і інші ДНК-глікозидази, що формує апіримідинові та апуринові ділянки в пошкодженій молекулі ДНК;
б) розщеплення ендонуклеазою 3',5'-фосфодіефірного зв’язку «зліва» від депіримідинізованого пентозофосфатного залишку;
в) вбудова «правильної» азотистої основи (в даному випадку — цитозину) на місце видаленої основи за рахунок дії ДНК-полімерази;
г) зшивання розриву в ланцюгу ДНК-лігазою.