Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ IV. МОЛЕКУЛЯРНІ МЕХАНІЗМИ СПАДКОВОСТІ ТА РЕАЛІЗАЦІЇ ГЕНЕТИЧНОЇ ІНФОРМАЦІЇ
ГЛАВА 22. РЕГУЛЯЦІЯ ЕКСПРЕСІЇ ГЕНІВ. ГЕНЕТИЧНІ РЕКОМБІНАЦІЇ
22.4. ГЕННА ІНЖЕНЕРІЯ. РЕКОМБІНАНТНІ ДНК
Рекомбінації генів, що були розглянуті вище, є природним процесом, який реалізується in vivo і є біологічним механізмом, спрямованим на збільшення різноманітності генетичного матеріалу в клітинах різних організмів.
Генна інженерія, або технологія рекомбінантних ДНК — науково-практичний напрямок сучасної біомедичної науки, основою методології якого є виділення з клітин індивідуальної ДНК та спрямоване маніпулювання з її молекулами, зокрема отримання молекулярних химер, тобто молекул, сформованих із фрагментів ДНК різних біологічних видів.
Біомедичне значення методів генної інженерії
Біотехнологічні методи генної інженерії мають на меті:
1) отримання нових генотипів (та фенотипів) організмів шляхом трансплантації гена одного організму в генотип іншого. Конкретними досягненнями цього біотехнологічного напрямку є створення химерних форм мікроорганізмів, що містять в собі гени, які спроможні спрямовувати синтез корисних для людини білкових продуктів, зокрема лікарських засобів (інтерферонів, гормонів (інсуліну тощо), ферментів, імунобіологічних препаратів (зокрема, вакцини проти вірусу гепатиту В) тощо;
2) застосування «генних трансплантацій» для лікування спадкових хвороб людини та тварин — генна терапія. Цей напрямок біотехнологічних досліджень знаходиться на початку свого розвитку і обіцяє в майбутньому створити принципово нові медичні технології терапії практично невиліковних в наш час спадкових хвороб та значної кількості найбільш поширених хвороб, у патогенезі яких певне місце займають спадкові фактори (атеросклероз, цукровий діабет, нервово-психічні хвороби тощо). Такий підхід був уже реалізований у терапії спадкового гіпогонадизму у мишей, в запліднену яйцеклітину яких був імплантований ген гонадоліберину.
Технологія трансплантації генів:
1) отримання в чистому вигляді, тобто у формі ізольованого фрагмента ДНК, гена з певними властивостями (тобто такого, що кодує синтез необхідного ферменту, гормону тощо);
2) конструювання рекомбінантної (гібридної, химерної) молекули ДНК;
3) введення рекомбінантної ДНК всередину реципієнтної бактеріальної клітини (тобто такої, в якій буде здійснюватися реплікація, клонування необхідного гена);
4) клонування рекобінантної ДНК.
1. Отримання необхідного гена.
Отримання гена (молекули ДНК), що буде підлягати реплікації (клонуванню) з виходом значної кількості реплік, може бути здійснено такими методами:
- хімічним синтезом гена (можливо тільки для коротких генів, що складаються з декількох десятків азотистих основ);
- виділенням необхідного гена (фрагмента ДНК) з цілісного геному клітини (проблема утруднена у зв’язку із значною складністю геномів еукаріотів, у складі яких значна кількість інтронів та повторів, які не транскрибуються — див. вище);
- конструюванням на мРНК (що кодує синтез білка, який бажано отримати в результаті біотехнологічної процедури) комплементарної відносно неї ДНК (кДНК). Цей метод потребує застосування зворотної транскриптази — ферменту, що присутній в деяких РНК-вмісних вірусах і забезпечує синтез ДНК на РНК матриці згідно з рівнянням реакції:
![]()
Метод широко використовується для отримання кДНК і включає в себе виділення з тотальної мРНК тканини мРНК, що кодує трансляцію певного білка (наприклад, інтерферону, інсуліну) з подальшим синтезом на цій мРНК як на матриці необхідної кДНК за допомогою зворотної транскриптази.
2. Конструювання рекомбінантної ДНК.
Ген, що був отриманий за допомогою вищерозглянутої процедури (кДНК), необхідно ввести в бактеріальну клітину таким чином, щоб він інтегрувався в її геном. Для цього формують рекомбінантну ДНК, що складається з кДНК та особливої молекули ДНК, яка править за провідник, або вектор, здатний до проникнення в реципієнтну клітину.
У ролі векторів для кДНК застосовують віруси або плазміди. Плазміди — це невеличкі кільцеві молекули ДНК, які розташовані окремо від нуклеоїду бактеріальної клітини (зокрема, E. Coli), містять у своєму складі декілька важливих для функції всієї клітини генів (наприклад, гени стійкості до антибіотиків) і можуть реплікуватися незалежно від основного геному (ДНК) клітини. Біологічно важливими і практично корисними для генної інженерії властивостями плазмід є їх здатність до переходу з однієї клітини в іншу за механізмом трансформації або кон’югації, а також спроможність включатися в бактеріальну хромосому та реплікуватися разом з нею.
Для конструювання рекомбінантної ДНК як кільцеву ДНК плазміди, так і лінійну кДНК розщеплюють за допомогою високоспецифічних до певних нуклеотидних послідовностей ендонуклеаз — так званих рестриктаз.
Рестриктази (рестрикційні ендонуклеази) — ферменти, присутність яких в бактеріальній клітині «обмежує» (restriction — обмеження, англ.) можливість розмноження в них певних бактеріофагів за рахунок розщеплення рестриктазами фагових ДНК. Назви рестриктаз ґрунтуються на назвах штамів бактерій, з яких їх виділяють: наприклад, рестриктази Eco RI та Eco RII походять з R-штаму E. Coli, рестриктаза Bam HI — з H-штаму мікроорганізму Bacillus amiloliquefaciens.
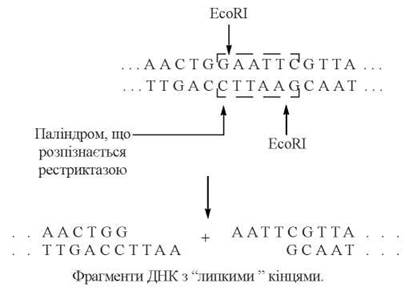
Рестриктази (яких на даний час відомо більше 200 типів) розщеплюють дволанцюгову ДНК лише в ділянках певних нуклеотидних послідовностей (звичайно, довжиною в 4-7 азотистих основ, які мають структуру паліндромів), що дозволяє використовувати ці ендонуклеази для «розрізання» молекул ДНК в суворо визначених сайтах. Наприклад, рестриктаза Eco RI гідролізує зв’язки між Gта A в таких послідовностях:
...GAATTC...
...CTTAAG...
Розщеплення рестриктазою G-A - зв’язків у зазначених сайтах дволанцюгової ДНК, як видно із наведеної нижче схеми, супроводжується утворенням в молекулі «кінців, що стирчать».
Одноланцюгові кінці молекули ДНК, «що стирчать», називать також «липкими» кінцями, оскільки вони є ділянками не- спарених нуклеотидів, що можуть легко сполучатися з комплементарними їм полінуклеотидними ланцюгами:
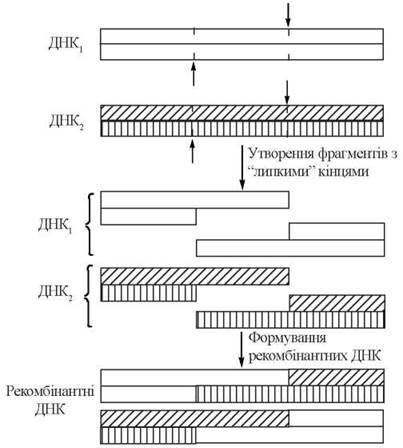
Розщеплення ДНК плазміди та кДНК (гена) специфічними рестриктазами призводить до утворення розрізаних в певних сайтах молекул ДНК з «липкими» кінцями. При взаємодії in vitro«розрізаних» плазміди та гена їх полшуклеотидт ланцюги, що складають «липкі» кінці, взаємодіють із утворенням водневих зв’язків між комплементарними основами, як це зображено вище. Застосування ДНК-лігази, яка утворює 3'-5'-фосфодіефірні зв’язки між кінцевими нуклеотидами, призводить до «зшивання» плазміди та гена і завершує утворення рекомбінантної ДНК (рис. 22.16):
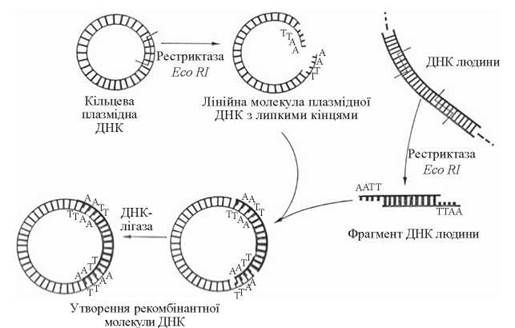
Рис. 22.16. Схема утворення рекомбінантної ДНК з плазміди та фрагмента ДНК людини.
3. Введення рекомбінантної ДНК всередину реципієнтної клітини та клонування необхідного гена.
Рекомбінантні ДНК, що складаються з плазмідної ДНК та ДНК гена, що трансплантується (наприклад, гена, який кодує синтез певного білка організму людини) при взаємодії з бактеріальними клітинами (що є нормальними хазяїнами для даної плазміди), можуть проникати всередину останніх.
Усередині клітини хазяїна відбувається реплікація (клонування) рекомбінантної ДНК з утворенням багатьох тисяч копій. У подальшому ці клоновані ДНК виходять з бактеріальної клітини, і з них можливо (знову ж таки за допомогою рестриктаз) виділити велику кількість копій шуканого гена.
У наш час для клонування генів, крім бактерій, використовують клітини дріжджів, грибів, рослин і навіть вищих тварин. За технологією, що розглянута, здійснено генно-інженерний синтез інтерферону людини, людських інсуліну, гормону росту, соматостатину, активатора плазміногену, білкових препаратів для діагностики СНІДу тощо.