Биохимия и молекулярная биология - Белясова Н.А. 2002
Метаболизм. Процессы, требующие притока энергии
Метаболизм азотсодержащих соединений
Выведение аммиака из организма
При деградации аминокислот, которая у млекопитающих осуществляется в печени, высвобождается аммиак. Кроме этого, значительные количества аммиака образуются при распаде пуринов и пиримидинов. Часть аммиака сразу расходуется на синтез других аминокислот и азотистых оснований, а оставшаяся часть должна быстро инактивироваться или выводиться из организма, поскольку это соединение является сильным клеточным ядом.
У организмов, стоящих на низших ступенях развития, а также у обитающих в воде животных аммиак выделяется непосредственно из клеток, а у рыб, например, через жабры. Такие животные носят название аммониотелические. У большинства наземных позвоночных животных, в том числе у млекопитающих и у человека, аммиак превращается в мочевину. Организмы, удаляющие аммиак (основную его часть) в виде мочевины называются уреотелические. Наконец, существуют урикотелические организмы (птицы и рептилии), которые превращают аммиак в мочевую кислоту и выводят ее из организма в твердом виде.
Свойства мочевины как вещества, в составе которого аммиачный азот выводится из уреотелических организмов, заключаются в том, что это нейтральное, нетоксичное, низкомолекулярное соединение, способное легко (в ходе пассивной диффузии) преодолевать мембранные барьеры, растворимое в воде. Мочевина легко переносится кровью и выводится с мочой.
Образуется мочевина в клетках печени в замкнутой последовательности реакций, которые носят название цикл мочевины.
Цикл мочевины. Один из атомов азота в составе молекулы мочевины происходит из аммиака, второй — из аспартата. Углеродный атом мочевины берет происхождение из молекулы СО2.
На первой стадии свободный аммиак и углекислота образуют в АТР-зависимой реакции молекулу карбамоилфосфата, которая содержит ангидридную связь и характеризуется высоким потенциалом переноса карбамоильной группы. Карбамоильный остаток переносится на орнитин с помощью орнитин-карбамоилтрансферазы и образуется цитруллин (рис. 16.13).
На следующей стадии при участии аргининосукцинат-синтазы в цикл включается вторая аминогруппа, которая присутствует в составе аспартата. Конденсация цитруллина с аспартатом также сопряжена с гидролизом АТР и приводит к образованию аргининосукцината.
На последнем этапе с участием аргининосукциназы происходит расщепление ее субстрата на аргинин и фумарат. Аргинин гидролизуется аргиназой на мочевину и орнитин, который вновь может акцептировать карбамоильный остаток. Фумарат может превращаться в оксалоацетат (реакции ЦТК), который в ходе трансаминирования с участием глутамата преобразуется в аспартат. Аспартат снова может поступать в цикл мочевины, конденсируясь с цитруллином (рис. 16.13).
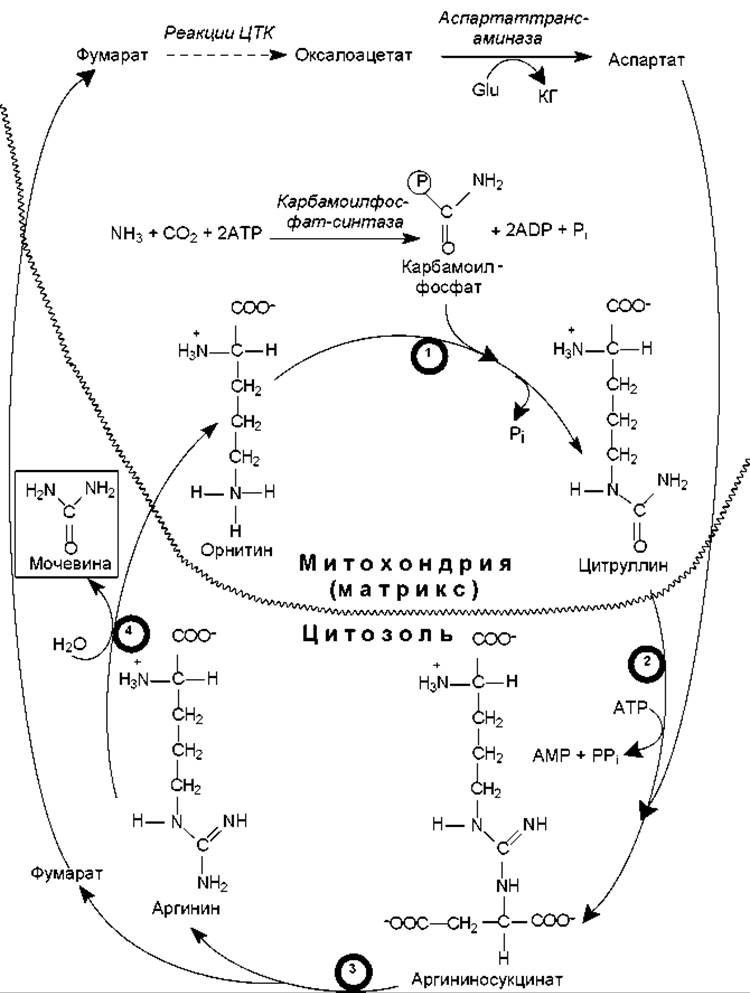
Рис. 16.13. Цикл мочевины. В кружках под стрелками — названия ферментов: 1 — орнитин-карбамоилтрансфераза; 2 — аргининосукцинат-синтаза; 3 — аргининосукцинат-лиаза; 4 — аргиназа. Обозначения: Glu — глутамат; КГ — a-кетоглутарат
Компартментализация цикла мочевины охватывает митохондрии и цитозоль: в матриксе протекают реакции образования карбамоилфосфата, его включение в состав цитруллина и преобразование фумарата в аспартат. Остальные три реакции (образование аргининосукцината, аргинина и фумарата, орнитина и мочевины) осуществляются в цитозоле.
На образование одной молекулы мочевины расходуется четыре макроэргических связи АТР, т. е. процесс инактивации аммиака стоит клетке больших затрат энергии.
Регуляция скорости цикла мочевины определяется первой реакцией. Катализирующая ее карбамоилфосфат-синтаза аллостерически активируется N-ацетилглутаматом, предшественником орнитина, и в его отсутствие практически неактивна.
Цикл мочевины — основной процесс, в котором происходит обезвреживание аммиака. Однако даже уреотелические организмы способны выделять небольшую долю аммиака непосредственно при гидролитическом дезамидировании глутамина и окислительном дезаминировании глутамата в клетках почек. Высвободившийся аммиак диффундирует через клеточные мембраны почечных трубочек в почечные канальцы (мочу), где соединяется с протонами, образуя ионы аммония. В такой форме аммиак уже не может преодолеть мембранный барьер и возвратиться в клетку, поэтому полностью выводится в составе мочи.
Известны наследственные заболевания, связанные с нарушением цикла мочевины из-за дефектов в его ферментной системе. Это всегда связано с увеличением концентрации аммиака в тканях и крови (гиперам-монемия), что может приводить даже к летальному исходу. При подобных заболеваниях страдают в первую очередь наиболее чувствительные к снижению содержания АТР системы — нервные клетки и мозг. Это можно объяснить тем, что высокая концентрация аммиака сдвигает равновесие обратимой реакции, катализируемой глутаматдегидрогеназой, в сторону образования глутамата, что неизбежно вызывает истощение запасов a-кетоглутарата. Но a-кетоглутарат является промежуточным продуктом ЦТК, и его чрезмерное потребление приводит к замедлению скорости цикла лимонной кислоты — основного поставщика восстановительных эквивалентов для дыхательной цепи. В результате уменьшается скорость образования АТР.