ЕКОЛОГІЧНА БІОХІМІЯ - Навчальний посібник - В. М. Ісаєнко 2005
Розділ 6. ЕКОЛОГО-БІОХІМІЧНА ВЗАЄМОДІЯ МІЖ ТВАРИНАМИ
6.1.Феромони безхребетних тварин
Серед феромонів безхребетних тварин вирізняють кілька груп залежно від їхніх функцій: 1) статеві (приваблювання особин протилежної статі, стимуляції спарювання, формування і зміни статевих ознак, стимуляції статевого дозрівання та розмноження); 2) занепокоєння; 3) агрегації (скупчення); 4) слідові; 5) мітки (мічення).
Зауважимо, що більшість феромонів є поліфункціональними — виконують кілька функцій, наприклад, приваблювання, мітки тощо.
Ці групи феромонів установлено для всіх безхребетних тварин, але найкраще вивчено для комах. Структуру деяких із них наведено нарис. 6.1.
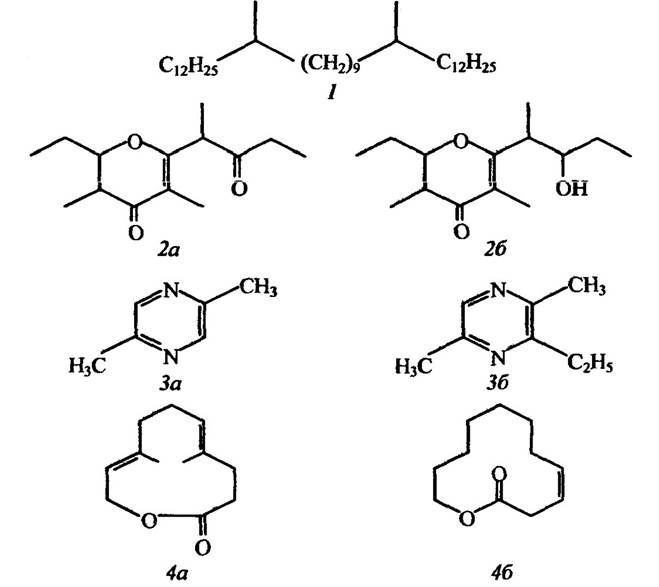
Рис. 6.1. Структура деяких феромонів комах:
1 — статевий феромон мухи цеце Glossina pallidipes; 2 — статеві феромони жука Lasioderma serricome: (2,3-цис)-серикорн (2а) і (2,3-цис)-серикорол (26); 3 — феромони сліду мурашок Tetramorium caespitum: 2,5-диметилпіразин (За) і 3-етил-2,5-диметилпіразин (3б); 4—агреаційний феромон самців лука Cryptolestes ferrugineus: ферулактон І(4а) і ферулактон 11(46).
Феромони водних безхребетних також різноманітні за функціями та структурою. Серед них існують статеві, стимулятори розвитку яєчників, інгібітори росту, стимулятори рухової активності та ін. Структуру деяких із них ілюструє рис. 6.2.
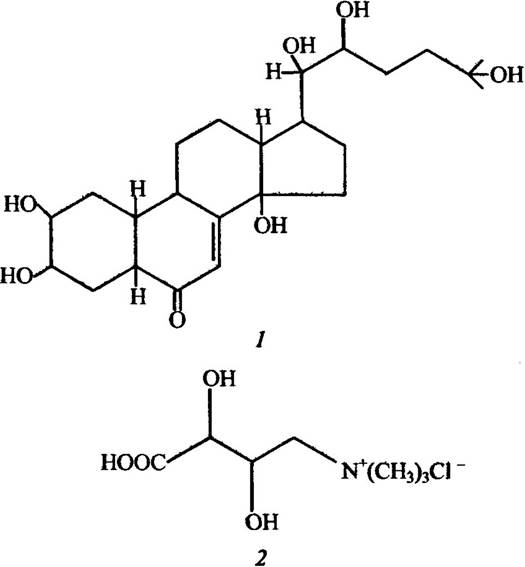
Рис. 6.2. Структура деяких феромонів морських безхребетних:
1 — статевий феромон крустекдизон деяких видів крабів; 2 — феромон занепокоєння антоплеурин актинії Anthopleura elegantissima
Статеві феромони приваблювання та стимуляції копуляції.
Ці феромони поділяють на дві групи: 1) статеві атрактанти — речовини, які сприяють зближенню самців і самок; 2) афродизиаки — речовини, що виділяються особинами в разі наближення потенційних статевих партнерів один до одного та сприяють підготовці особини іншої статі до копуляції.
Існує також інше трактування груп статевих феромонів. Так, під терміном «статеві феромони» інколи розуміють феромони самок для принадження та спонукання до спарювання самців, які перебувають на деякій відстані від них. Цей самий термін застосовують до феромонів, які виділяються самцями для спонукання до спарювання самок. Останні феромони інколи називають афродизиаками. Узагалі термін «афродизиаки» некоректний, якщо йдеться про статеві феромони. Він стосується лікарських препаратів, які посилюють лібідо в людей. Ми користуватимемо прийнятним першим визначенням поняття «статеві феромони».
Найпростішим за хімічною будовою є, очевидно, статевий атрактант валеріанова кислота — жіночий феромон лускунчика каліфорнійського. Однак більшість феромонів — ненасичені спирти, ацетати, карбоксилати тощо (табл. 6.2). Серед цих сполук є 9-кетодеценова кислота, яка виділяється маткою бджіл для принадження та спонукання трутнів до спарювання з нею.
Таблиця 6.2
СТРУКТУРА ДЕЯКИХ СТАТЕВИХ ФЕРОМОНІВ КОМАХ
(за Харборном, 1985, зі змін.)
Сполука |
Організм |
Валеріанова кислота СН3(СН2)3СО2Н |
Личинки лускунчика каліфорнійського Limonius californicus |
Транс-9-кето-2-деценова кислота СН3СО(СН2)5СН = СHCO2H |
Бджола медоносна Apis mellifera |
Цис-7-Додеценілацетат СН3(СН2)3СН = СН(СН2)6ОАц |
Совка Trichoplusia |
Цис-8-Додецепілацетат СН3(СН2)2СН = СН(СН2)7ОАц |
Східна плодожерка Grapholitha molesta |
Цис-11-Тетрадеценілацетат СН3СН2СН = СН(СН2)10ОАц |
Дубова листовійка Archips semiferanus |
Гексадеканілацетат СНз(СН2)15ОАц |
Метелик Lycorea ceres |
Примітка: Ац — ацетил.
Іноді статеві феромони мають у своєму складі циклічні структури, наприклад серикорн і серикорол у жука Lasioderma serricome (рис. 6.1), феромон довгоносика Anthonomust grandis — похідна циклогексану, а молі Leucania impurias — бензальдегід.
Досить докладно статеві феромони вивчено в шовкопряда Bombyx mori. Самки цього виду виділяють пахучими залозами леткий секрет бомбікол (гексадиен-10,12-ол-1) (рис. 6.3), який приваблює самців. Вони починають швидко трясти крилами та пророблювати характерні обертальні рухи. Це, очевидно, сприяє швидкому надходженню статевого феромону самця до самки, що стимулює її до копуляції.
Нині ідентифіковано й вивчено велику кількість статевих феромонів безхребетних.
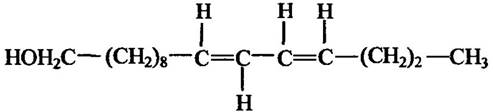
Рис. 6.3. Статевий феромон бомбікол самок шовкопряда Bombyx mori
Деякі природні та синтетичні сполуки здатні імітувати дію природних феромонів. Так, з екстрактів рослинних тканин голонасінних виділено D-борнилацетат, який викликає статеве збудження таргана американського Регіїапеіа атегісапа, синтетична сполука тримедлур активна стосовно середньоморської дрозофіли, а куелур — динної мухи Dacus cucurbitae (рис. 6.4).
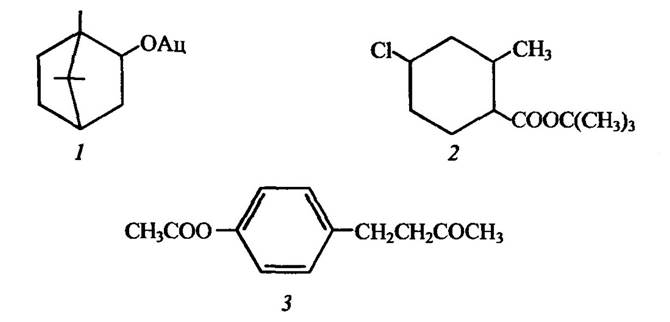
Рис. 6.4. Структура деяких сполук, які імітують статеві гормони комах:
1 — D-борнилацетат; 2 — тримедлур; 3 — куелур; Ац — ацетил
У деяких випадках під час вивчення статевої поведінки комах було виявлено специфічні взаємодії між комахами та рослинами. Так, Anthera polyphemus здатні спарюватися за наявності листя північного дуба Quercus borealis. Ця рослина з листя виділяє транс-2- гексаналь, який спричиняє виділення у самок феромону, що спонукає самців до спарювання.
Статеві феромони знайдено в нематод, які паразитують на рослинах. Вони беруть участь не тільки в регулюванні процесу спарювання, а й у попередженні спарювання нематод різних видів.
Феромони — модифікатори статі. Ці феромони теж належать до статевих. Проте на відміну від розглянутих вище статевих феромонів, основна функція яких — приваблювання особин іншої статі та заохочування до копуляції, ці здатні формувати або змінювати і ознаки статі. Представником таких феромонів є бонелін самок морського черва Bonellia viridis. Доросла самка цього черва має відносно товсте тіло завдовжки приблизно 7 см і тонкий роздвоєний на вершині хобот довжиною понад 90 см. Самка стає статевозрілою на другому році життя і продовжує рости ще кілька років. Самець досягає довжини лише 1 — 3 мм і живе в статевих протоках або органах виділення (нефридіях) самки. Якщо личинки самців розвиваються у статевонезрілої самки, 87 % з них перетворюються в самок і тільки 13 % — у самців. Якщо личинки самців помістити у воду, в якій знаходиться статевозріла самка, то майже всі вони прикріпляються до хобота самки і протягом трьох діб підпадають під дію феромону. Потім личинки самців відокремлюються та перетворюються в самців.
Утворення феромонів, які модифікують стать, виявлено в черевоногих молюсків роду Crepidula. Для цих гермафродитних молюсків характерна протинандрія — молоді особини спочатку проходять функціональну стадію самців, а потім — самок. Лише невелика кількість молодих особин перетворюється в самок без попереднього проходження стадії самців. Самки виділяють у воду феромон, який викликає швидкий перехід молодих особин у стадію самця та тривалу затримку на цій стадії розвитку.
Статевозрілі особини суспільних комах (термітів, мурашок, бджіл) продукують і виділяють речовини, які пригнічують розвиток статевих залоз у інших особин тієї самої родини.
У термітів (зокрема, Kalotermes flavicollis) робочі особини, які принаджуються феромонами самки й самця, віддають їм їжу і злизують виділення з покривів черевця й анальних отворів. Ці виділення містять феромони, які гальмують розвиток статевих органів і перетворення личинок і німф у статевозрілі особини. Сам по собі феромон самця істотно не впливає на личинок і німф, а самки — дещо гальмує їх перетворення в статевозрілих самок. Однак за одночасної дії феромонів самця та самки перетворення личинок і німф попереджується цілком.
У деяких видів термітів, наприклад Zootermopsis angusticolis, особини певних каст виділяють феромони, які перешкоджають перетворенню личинок на комах даної касти.
Яйцеродні самки мурашок також виробляють феромони, які принаджують робочих мурашок і пригнічують у них розвиток яєчників. Робочі мурашки злизують цю речовину з поверхні хітинового покриву самки та передають, відригуючи їжу, іншим особинам.
Для нормального функціонування колонії бджіл велике значення має поліфункціональний хеморегулятор, який продукується бджолиною маткою і називається «царською» речовиною. Ця речовина складається не менш як з 32 речовин, однією з яких є 9-кетодеценова кислота. Вона приваблює до матки робочих бджіл, які годують її. Облизуючи тіло матки, вони отримують «царську» речовину, дією якої є пригнічення розвитку яєчників і бажання будувати «царські» комірки для годування майбутньої матки. Крім цього, вона має статевий феромон, який приваблює самців до матки.
Феромони — стимулятори статевого дозрівання та розмноження. Ці феромони викликають в інших особин свого виду дозрівання статевих залоз і виділення гамет, приваблюють особин різної статі до спарювання, а в живородних тварин впливають на настання й перебіг вагітності.
Багатощетинкові черви родини Nereidae живуть на дні океану, але для розмноження піднімаються в поверхневі шари води. Коли настає час розмноження, тіло цих черв’яків ділиться впоперек на дві частини. Передня частина залишається на дні, а задня, яка містить статеві залози, піднімається до поверхні води. Мозок цих червів виробляє феромон, що гальмує дозрівання статевих залоз. Після роздвоєння мозок залишається в передній частині, а в задній — після відновлення головної частини гальмуючий феромон не виробляється і статеві клітини швидко дозрівають. Статевозрілі самки Nereis виділяють феромон, який стимулює самців до викидання у воду зрілих сперматозоїдів.
Статевозрілі самці поліхети Grubea clavata разом зі спермою викидають у воду феромон, який стимулює відкладання яєць у самок. Без цієї речовини дозрілі яйця залишаються в порожнині тіла самки, де швидко гинуть.
Статевозрілі самці сарани Schistocerca gregaria виділяють феромон, який значно прискорює статеве дозрівання інших самців і самок. Однак дія феромону тільки на нюхові рецептори є недостатньою для прискорення статевого дозрівання. З’ясовано, що для цього необхідний більш тісний взаємозв’язок статевонезрілих особин зі статевозрілими, який з’являється при утворенні щільних скупчень сарани.
Самці павуків Schizocosa ocreata виділяють феромон, що пригнічує характерну поведінку («залицяння») інших самців із павутиною самок.
У великої кількості видів комах самці виробляють речовини, які самки злизують перед спарюванням або під час нього. Ці речовини викликають у самок статеве збудження або малорухомий стан, який полегшує статевий акт. Так, наприклад, на різних ділянках тіла, найчастіше в заглибленнях лоба або на вершинах надкрил, самці жуків родини Malyridae мають залозисті органи, які називають ексцита-
торами. Перед спарюванням самець підставляє самці ті ділянки свого тіла, де розміщені ексцитатори. Самка захоплює щелепами покрови самця, доторкається до ексцитаторів вершинами щелеп і губних щупалець, де розміщені смакові рецептори. Отримані при цьому хімічні та механічні подразнення, очевидно, стимулюють самок до спарювання.
Феромони занепокоєння (тривоги), втечі, активної оборони.
Ці феромони виявлено у бджіл, мурашок, термітів, попелиць, блощиць та інших тварин. Деякі феромони занепокоєння є одночасно також захисними речовинами (рис. 6.5).
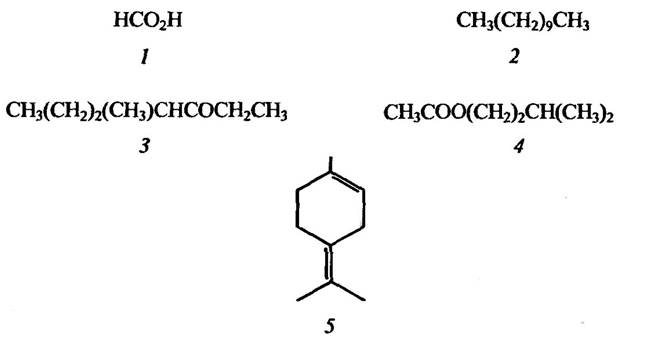
Рис. 6.5. Структура деяких феромонів занепокоєння:
1 — мурашина кислота мурашок Formica; 2 — ундекан мурашок Formica; 3 — 4-метил-З-гептанон мурашок Pogonomyrmex; 4 — ізоамілацетат бджіл Apis; 5 — терленолен термітів Armitermes
Жалючий апарат бджіл і ос містить кілька залоз, які продукують феромони занепокоєння. Сама отруйна залоза виділяє феромон занепокоєння, який виділяється разом з отрутою. Оси роду Vespa вприскують отруту разом із феромоном занепокоєння, а медоносні робочі бджоли Apis mellifera, ужаливши, залишають мітку (виділяє ізоамілацетат), що вказує іншим бджолам місце, яке треба жалити.
Зазвичай бджола не може витягнути своє жало зі шкіри хребетних тварин, оскільки воно має 12 зубчиків, спрямованих вістрям назад. Жало залишається у шкірі ворога й відривається разом із жалючим апаратом та його залозами від тіла бджоли. Хоча бджола після цього гине, отрута продовжує надходити в тіло ворога з резервуара отруйної залози. Крім цього, ворог, якого вжалила бджола, мітиться феромоном занепокоєння і стає, навіть при втечі, об’єктом нападу інших бджіл, які знаходять його запахи.
Феромони занепокоєння в комах мають просту структуру (рис. 6.5). У деяких мурашок це прості вуглеводи, такі як ундекан, тридекан і пентадеканол, альдегідні та кетонні похідні цих вуглеводів. Феромони занепокоєння в родин Formicinae, Hymenoptera, Isoptera та деяких інших становлять собою компоненти етерних олій, таких як цитронелона, цитраль, α-пінен, лімонен, терпинолен. У попелиць феромони занепокоєння, як правило, за хімічною структурою терпеноїди, зокрема моноциклічний сесквитерпен гермаркрен А.
Феромони занепокоєння виявлено також у попелиць. Якщо попелиці загрожує небезпека, то вона виділяє рідину, яка містить транс-β-фернезен, запах якого викликає реакцію «втечі» в різних видів попелиць.
Феромони занепокоєння найменше специфічні з летких гормонів комах. Так, 2-гексаналь є активним компонентом феромонів занепокоєння червоного клопа Dysdercus intermedius і мурашок Cremato- gaster africana. Однак феромони занепокоєння в різних організмів, як правило, неоднакові.
Кількість інформації, яка передається феромонами занепокоєння, може бути значною. Так, наприклад, мурашки Oecophiila longinoda в разі нападу хижака виділяють чотири феромони з різною летучістю. Найлеткішими є 1-гексаналь, який мурашки сприймають біля місця атаки. Наближаючись до нього, вони сприймають близький за структурою до першого феромону спирт 1-гексанол. Потім напад на хижака стимулюється 2-бутил-2-октенолом. Ще один, останній феромон, 3-ундеканон разом із третім, орієнтує мурашок на близькій відстані від хижака.
Мурашки самі здатні нападати на інших тварин і при цьому завдяки феромонам проявляють добре виражені взаємодії.
Феромони занепокоєння виявлено й у гідробіонтів. Так, при ушкодженні актиній Anthopleura elegantissima виділяється бетаїн (3-карбоксси-2,3-диокси-N,N,N-триметил)-1-пропанамоній, який спричиняє замикання щупалець і передає сигнал, що сприймається сусідніми організмами.
Феромони слідів і мітки. Ці феромони тварини наносять на ґрунт або навколишні предмети й використовують як орієнтири на території під час добування їжі та для інших видів активної діяльності. Особливе значення ці феромони мають для суспільних комах — мурашок, бджіл, термітів. За їхньою допомогою утворюються пахучі сліди від гнізда мешкання до джерела їжі. Від них залежить специфічний запах кожного мурашника, термітника, бджолиного вулика. Запах пахучого слідового феромону специфічний для кожного виду. Так, наприклад, мурашки-листорізи виробляють 4-метил-пірол-2-карбонову кислоту, яка сприймається мурашками в кількості 0,8 пг/см.
У мурашок Monomorium pharaonis виявлено слідовий феромон біциклічний алкалоїд.
Структуру деяких слідових феромонів наведено на рис. 6.6.
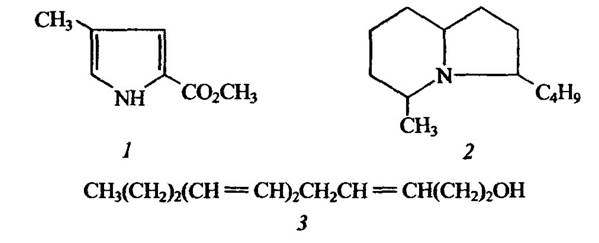
Рис. 6.6. Структура деяких слідових феромонів
1 — метиленовий етер 4-метилпірол-2-карбонової кислоти мурашок- листорізів Alta lexana;
2 — біциклічний алкалоїд фараонових мурашок Monomotium pharaonis;
3 — 3-цис- 6-трднс-8-додекатриен-1-ол термітів Reticulitermes virginicus
Сліди багатьох видів мурашок стимулюють робочих особин слідувати вздовж них. Однак деякі види мурашок наносять свої сліди у вигляді смужок або плям, які додають мурашкам можливість визначити напрямок, в якому ці сліди було прокладено.
Інтенсивність пахучого сліду, який нанесли мурашки, є показником кількості їжі в тому місці, куди веде слід. Мурашки, які повертаються в мурашник, наносять пахучі сліди тільки тоді, коли вони здобули їжу. Що більше їжі в певному місці, то більша кількість мурашок наносить пахучі сліди на зворотному шляху, то інтенсивні- ший запах сліду. З тим, як запаси їжі вичерпуються, зменшується кількість мурашок, які відвідують джерело їжі. Мурашки швидко перестають сприймати старі запахи, що вивітрюються, і їхній рух до джерела їжі, яке вичерпалося, припиняється.
Бджола, котра знайшла їжу, сама вказує шлях до неї іншим робочим бджолам своєї родини. Ще у вулику вона у вигляді характерного «танцю» сповіщає напрям польоту до джерела їжі й відстань до нього, а відтак, як правило, супроводжує робочих бджіл до джерела їжі. Проте одного зорового орієнтира для цього недостатньо. Тому бджола, яка знайшла джерело їжі, час від часу наносить слідовий феромон на стебла трави, гілки, каміння або інші предмети, які слугують орієнтиром. Що ближче до джерела їжі, то більше слідового феромону наносить бджола на поверхню орієнтирів.
Медоносна бджола Apis mellifera як слідовий феромон наносить монотерпен герані, який безпосередньо отримує з рослин і концентрує в організмі. Бджоли родини Trigona використовують інший слідовий феромон — бензальдегід, який може утворюватися з циано-генного глікозиду прунасину.
Подібно до бджіл чинять і джмелі. Так, наприклад, із гнізда джмелів Bombus hortorum вранці першими вилітають самці, які на різні предмети біля гнізда та вздовж шляху польоту наносять слідовий феромон фарнесон (рис. 6.7).
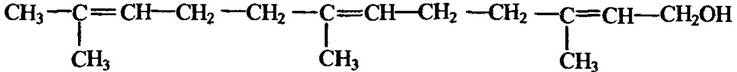
Рис. 6.7. Структура феромону фарнесон джмелів
Терміти Reticulitermes virginicus як слідовий феромон використовують 3-г/мс-6-/иршс-8-додекатриен-1 -ол, що виявлено також у грибах деревини, якою харчуються терміти. Капронова кислота є слідовим феромоном у термітів Zootermopsis nevadensis.
Слідові феромони суспільних комах можуть використовуватися хижаками (жуками, кліщами, багатоніжками, зміями та ін.), які відшукують за їхнім запахом гніздомешкання.
Феромони мітки (мічення) близькі до слідових феромонів, але мають відмітну функцію — слугують пахучими мітками, які вказують на зайнятість даної території певною особиною або родиною. Такі феромони притаманні багатьом комахам, у тому числі хижим. Ці комахи мітять, зокрема, такими феромонами обстежену ними територію, що збільшує ефективність пошуку жертв і скорочує на це час.
Різновид таких феромонів — феромони овіпозиції (відкладання яєць) (див. розд. 4). Ними самка, яка відкладає яйце, мітить саме яйце або місце його відкладання. Це дає можливість уникнути повторного відкладання яйця в це саме місце і, отже, вилуплення личинок і труднощів з кормовою базою. У цьому разі зменшується конкурсиція та ефективніше використовується корм.
Самки комах-паразитів під час своєї яйцекладки мітять ті яйця господаря, які вони заразили. Речовина, якою помітили яйця господаря, відлякує інших самок паразитуючої комахи.
Агрегаційні (скупчення) феромони. Ці феромони приваблюють особин свого виду. Вони описані в перетинчастокрилих, двокрилих, метеликів, жуків, тарганів, термітів, клопів і представників інших рядів. Нерідко ці речовини приваблюють особин обох статей до міс
ця живлення й розмноження. Завдяки цьому в певному місці можуть утворюватися великі скупчення комах, які належать до одного виду. Це значно полегшує зустріч самців і самок. Типовим прикладом дії таких феромонів є скупчення клопів-черепашок Euiygaster integriceps, небезпечного сільськогосподарського шкідника. Характерний запах постільних клопів Cimex lectularius приваблює інших особин цих ектопаразитів у певні кутки та тріщини стін, під плінтуси і шпалери, під розвішані на стінах килими й картини тощо.
Кімнатні мухи Musca domestica, коли сідають на їжу, виділяють летку речовину « мушиний фактор», яка приваблює інших мух цього виду. Тому муха, яка сіла на їжу, принаджує велику кількість інших.
Таргани Blattella germanica виділяють речовину, запах якої сприяє скупченню тарганів цього виду в певних місцях.
Особливо докладно вивчено феромони агрегації жуків-короїдів і лубоїдів родів Ips, Scolytus, Xyleborus і Dendroctonus. Самці Ips confusus, потрапивши у флоему та камбій дерева, виділяють феромон, який приваблює самців і самок цього виду. У лубоїдів Dendroctonus аналогічний феромон виділяють самки, а не самці.
Агрегаційні феромони можуть бути використані самцями або самками для приваблювання особин іншої статі. Тому ці феромони не завжди можна відрізнити від статевих феромонів, особливо в жосткокрилих. Водночас агрегаційні феромони можуть бути репелентами для особин тієї самої статі. Так, наприклад, у Dentroctonus adjunctus феромон бревікомін, який виділяється самцями, діє на самок як агрегаційний, а на інших самців — як репелент.
Самці Dendroctonus pseudotsugae виділяють комбінацію феромонів франталину та 3-метил-2-циклогексен-1-ону, яка діє на інших самців і самок цього виду як репелент. Якщо ж ці дві речовини поєднані з третьою — 3-метил-циклогексен-1-олом, то ця суміш діє як агрегаційний феромон для самців і самок.
Зауважимо, що агрегаційний феромон, як правило, становить собою комплекс феромонів, який охоплює також леткі речовини дере- ва-господаря.
Один зі способів використання феромонів для боротьби зі шкідниками — це сумісне використання статевих феромонів з інсектицидами, що дає можливість локалізувати місця внесення останніх. Використання феромонів занепокоєння з інсектицидами дає змогу значно зменшити дози інсектицидів, оскільки феромони занепокоєння викликають підвищення рухливості комах, а це призводить до збільшення ймовірності контакту з інсектицидом.
У цілому феромони та їхні аналоги або імітатори дії є одним з основних заходів біологічної боротьби зі шкідниками. їх застосу
вання сприяє значному зменшенню використання токсичних для тварин і людини хімічних сполук.
Перспективним є також використання феромонів гідробіонтів для оптимальної організації аквакультури.