ЕКОЛОГІЧНА БІОХІМІЯ - Навчальний посібник - В. М. Ісаєнко 2005
Розділ 8. СТІЙКІСТЬ ТА АДАПТАЦІЯ РОСЛИН І ТВАРИН
8.3. Стійкість та адаптація тварин до умов існування
8.3.1. Дихання тварин у воді та повітрі
Тварини використовують для енергозабезпечення життєдіяльності енергію хімічних сполук їжі. Більшість тварин отримує енергію шляхом аеробного (за участю оксигену) окиснення цих сполук. Кінцевими продуктами повного окиснення основних компонентів їжі, а саме вуглеводів, жирів і білків, є Н2О та СО2г. За окиснення білків утворюється ще деяка кількість інших кінцевих продуктів.
Деякі безхребетні тварини, як і певні бактерії, здатні використовувати енергію органічних сполук в анаеробних (безоксигенних) умовах. Однак в остаточному підсумку виділення енергії за цих умов становить не більше 10 % енергії, яка б утворилася за повного окиснення цих сполук. Зауважимо, що аеробному етапу розщеплення вуглеводів незалежно від наявності оксигену передує етап анаеробного перетворення.
Процес поглинання оксигену та виділення СО2 організмами називають диханням. Тварини вилучають оксиген із середовища, в якому вони живуть. У це саме середовище вони виділяють СО2. Утворену в процесі окиснення воду тварини залучають до загальної кількості води організму. Водні тварини використовують оксиген, який розчинений у воді, а деякі з них, зокрема двоякодихаючі риби, — також оксиген повітря. Наземні ж тварини дихають оксигеном, який міститься в атмосферному повітрі. Деякі тварини здатні отримувати достатню кількість оксигену через поверхню тіла, але більшість тварин мають спеціальні органи дихання. Основним, а в деяких випадках і єдиним процесом, який забезпечує потраплення оксигену із зовнішнього середовища до клітин, є дифузія, тобто переміщення оксигену в напрямі від великих концентрацій до низьких. Виведення СО2 із клітин відбувається також за концентраційним градієнтом — від більших концентрацій до менших. У тварин, які мають систему кровообігу, дифузії оксигену сприяє циркуляція крові.
Вміст оксигену в повітрі і воді. Склад атмосферного повітря у звичайних умовах постійний. Конвекційні потоки забезпечують його інтенсивне перемішування. Певне збагачення верхніх шарів атмосфери на висотах близько 100 км гідрогеном і гелієм не істотне для біосфери. Відносний склад сухого атмосферного повітря наведено в табл. 8.2.
Склад атмосферного повітря підтримується рівновагою між використанням оксигену в окисних процесах організмів та його вивільненням під час фотосинтезу (фоторозщепленням Н2О). Побоювання, що спалювання викопного палива (вугілля, нафти, природного газу) призведе до зменшення вмісту оксигену в атмосферному
повітрі, очевидно, безпідставні. Розраховано, що коли всі відомі запаси викопного палива буде спалено, то вміст оксигену в атмосфері зменшиться не більш як на 20,8 %.
Таблиця 8.2
ВІДНОСНИЙ СКЛАД СУХОГО АТМОСФЕРНОГО ПОВІТРЯ
Компонент повітря |
Вміст, % |
Оксиген (О2) Діоксид карбону (СО2) Нітроген (N2) Інертні гази (аргон, гелій, неон, криптон, ксенон) |
20,95 0,03 78,09 приблизно 1 |
Збільшення в атмосферному повітрі СО2 унаслідок спалювання викопного палива також не має викликати впливу на біоту. Разом з тим навіть незначне підвищення вмісту СО2 в атмосфері може впливати на поглинання енергії Сонця повітрям і викликати так званий парниковий ефект, здатний змінити кліматичні умови на Землі. Як відомо, атмосфера більш проникна для короткохвильового випромінювання, яке потрапляє ззовні, ніж довгохвильового, що випромінюється Землею. Довгохвильове випромінювання Землі поглинається головним чином СО2 атмосфери та водяною парою. Розраховано, що підвищення вмісту СО2 в атмосфері вдвічі призведе до збільшення її температури приблизно на 1,3 °С, якщо кількість водяної пари залишиться незмінною. Водночас підвищення температури атмосфери сприяє утриманню водяної пари, що спричинить утворення хмар, які відбиватимуть сонячне випромінювання, і це призведе до зменшення кількості енергії, що надходить на Землю. Разом з тим збільшення в атмосфері водяної пари призведе до зменшення кількості енергії довгохвильового випромінювання, яке випромінюється Землею й надходить у космічний простір, подібно до впливу підвищеного вмісту СО2. Отже, можливе виникнення парникового ефекту від підвищення концентрації СО2 є неоднозначним і потребує докладного дослідження.
Зауважимо, що сталість складу атмосфери спостерігається в повітрі над Землею. У ґрунті часто міститься мало оксигену через наявність у ньому органічних сполук, які окиснюються, або неорганічних, що поглинають оксиген (наприклад, сульфіду феруму). Такі процеси залежать від вмісту цих речовин, температури, вологості та інших факторів. Вміст оксигену може зменшуватися, а СО2 — збільшуватися в ущелинах, печерах, норах тощо.
Істотне значення атмосферний тиск має для збереження абсолютної кількості газу в атмосферному повітрі. Так, на рівні моря, де атмосферний тиск дорівнює 101,3 кПа (1 атм, 760 мм ртутного стовпчика) парціальний тиск (частка загального тиску газів, яка припадає на даний газ) оксигену в сухому повітрі становить 21,2 кПа (20,95 %). На висоті 6000 м атмосферний тиск оксигену буде 10,6 кПа (ті ж 20,95 %), але для життя людини він недостатній.
Кількість газів, яка розчиняється у воді, залежить від природи газу, його тиску в атмосфері, температури, наявності інших розчинних речовин тощо. Перехід газу у воду відбуватиметься доти, доки не встановиться рівновага, за якої однакова кількість молекул газу входитиме у воду та виходитиме з неї.
Розчинність газів у воді різна. Для ЇЇ характеристики введено поняття коефіцієнта розчинності — об’єм газу в мілілітрах, який розчиняється віл води за умов, коли тиск даного газу без водяної пари дорівнює 101,3 кПа, а температура—0 °С. Як правило, для біологічних досліджень розчинність газів наводять за температури 15 °С. Розчинність у воді основних газів атмосфери наведено в табл. 8.3.
Таблиця 8.3
КІЛЬКІСТЬ ОКСИГЕНУ, НІТРОГЕНУ Й ДІОКСИДУ КАРБОНУ У ВОДІ ЗА 15 °С І ТИСКУ 101,3 КПА
(за Шмідтом-Ніельсоном, 1982)
Назва газу |
Розчинність газу віл води |
Оксиген (О2) Нітроген (N2) Діоксид карбону (СО2) |
34,1 16,9 1019,0 |
Примітка. Вода перебуває в рівновазі з чистим безводним газом, який розчиняється в ній
Розчинність у воді діоксиду карбону приблизно в 30 разів більша, ніж оксигену, і в 60 разів більша, ніж нітрогену.
Кожний газ у суміші з іншими газами розчиняється залежно від власного парціального тиску і незалежно від наявності інших газів. Якщо парціальний тиск газу змінюється, то змінюється також кількість розчинного газу у воді: зі зменшенням тиску розчинений газ виходить із води до досягнення стану рівноваги та навпаки — з підвищенням тиску його розчинена кількість збільшується.
Розчинність газів зменшується з підвищенням температури і збільшенням кількості розчинених у ній інших речовин. У табл. 8.4 наведено відомості стосовно розчинності оксигену за різних температур у прісній і морській воді.
Таблиця 8.4
КІЛЬКІСТЬ РОЗЧИНЕНОГО У ПРІСНІЙ І МОРСЬКІЙ ВОДІ ОКСИГЕНУ
(за Шмідтом-Ніельсоном, 1982)
Температура, °С |
Прісна вода, мл газу в 1л води |
Морська вода, мл газу віл води |
0 |
10,29 |
7,97 |
10 |
8,02 |
6,35 |
15 |
7,22 |
5,79 |
20 |
6,57 |
5,31 |
30 |
5,57 |
4,46 |
Примітка. Вода перебуває в рівновазі з атмосферним повітрям.
Оскільки розчинний у воді газ перебуває в рівновазі зі своєю газовою фазою за даного парціального тиску, то газ у воді має такий самий парціальний тиск.
Дихання у воді. Водні тварини мають різні пристосування для дихання. Більшість дрібних тварин отримують оксиген із води шляхом його дифузії через поверхню тіла й не мають спеціальних органів дихання. Добре аерована вода перебуває в рівновазі з атмосферою, і тиск 02 у ній дорівнює 21,27 кПа. Водночас у дрібних організмів, які мають сферичну форму діаметром 1 мм, оксиген досягне центру сфери дифузією за умови його тиску біля поверхні не менше 15,2 кПа. Розрахунки велися за рівнянням Харві:
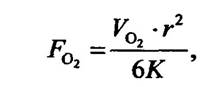
де FО2 — концентрація оксигену біля поверхні сфери порівняно з атмосферною; V О2— споживання оксигену в мілілітрах на 1 г за хвилину (0,001 мл/хв • г); K — константа дифузії, яка вказує на кількість мілілітрів O2, що дифундує за 1 хв через площу 1 см2 при градієнті O2 101,3 кПа/см (для сполучної та багатьох інших тканин тварин дорівнює 1,086 • 10-7 см2 • Па/хв); г— радіус сфери в сантиметрах.
Отже, дрібні організми (наприклад, найпростіші, деякі плоскі черви) з діаметром 1 мм і менше здатні шляхом дифузії споживати оксиген. Цим самим способом можуть споживати оксиген організми, що мають дуже низький рівень метаболізму (малі потреби оксигену), наприклад медузи. Хоча медузи можуть мати великі розміри, але вони містять, як правило, менше 1 % органічних сполук, а інші сполуки — це вода й солі. У медуз, крім того, метаболічно активні клітини наявні біля поверхні, де шлях дифузії оксигену відносно короткий.
В організмів, які мають несферичну форму, відносна поверхня тіла більша і коротший шлях дифузії. Ці організми, які мають сплющену або ниткоподібну форму, утворюють псевдоподії або складнішу поверхню. Прикладом таких організмів можуть бути корали, губки. Завдяки цьому вони здатні отримувати достатню кількість оксигену шляхом дифузії.
Якщо дихальна поверхня вивернута назовні й утворює випинання, то її зазвичай називають зябрами. Вторинно зябра можуть міститися в порожнині, як це спостерігається, наприклад, у риб.
Зябра мають тонкі стінки, що полегшує газообмін. Однак це призводить до зменшення механічної жорсткості. Тому в повітрі зябра не витримують особистої ваги і злипаються. Саме тому риби, витягнуті з води, страждають від задухи, хоча оксигену в повітрі більше, ніж у воді.
Для постачання оксигеном водних організмів важливе створення току води над поверхнею зябер. Для цього необхідно, щоб зябра рухались або переміщувалася сама вода. Рух зябер спостерігається в невеликих організмів, наприклад, личинок одноденок Ephemeroptera. За руху зябер необхідно збільшувати їхню механічну міцність, а це призводить до збільшення їхньої товщини, що зменшує здатність до газообміну. Однак велика водна амфібія Necturus переміщує свої зябра, але ці рухи повільні.
Рух води над поверхнею зябер забезпечується рухом вій (наприклад, у двостулкових молюсків), прогоном води через отвори за допомогою джгутиків (губки) або пристосуваннями, подібними до насоса (краби, риби). У деяких водних тварин руху води навколо поверхні зябер сприяє переміщення всього тіла. Типовим прикладом є безперервний рух тунця. Кальмари й восьминоги набирають воду в мантійну порожнину та викидають її через сифон.
Зябра крім дихальної функції виконують також інші, зокрема осморегуляцію (риби, краби та ін.), затримують зважені частини їжі і спрямовують їх у рот (вії двостулкових молюсків).
Зябра містяться в зябровій порожнині, яка забезпечує захист цих органів і дозволяє воді обтікати зяброву поверхню найефективнішим способом. Особливу перевагу надає така будова зябер, коли потік води над їхньою поверхнею і потік крові в зябрах спрямовані протилежно один одному (протитік). Це дає змогу насичуватися оксигеном крові до рівня, який існує у вхідній воді. Якби вода і кров текли в одному напрямі, то в кращому разі кількість оксигену в крові могла досягнути того самого рівня, що й у воді, яка витікає.
Існує кілька пристосувань у водних організмів для збільшення току води через зябра. Так, наприклад, кісткові риби використовують комбіновану дію ротової порожнини та зябрових кришок за участі відповідних клапанів для контролю потоку води. У деяких швидкоплавних риб (наприклад, тунців) вода безперервно тече через зябра (таранна вентиляція).
Для вилучення твердих частинок, які застряють у зябрах під час протікання води через них, водні організми змінюють напрям потоку води. Для цього вони розширюють ротову порожнину при зімкнутих губах. Це викликає зниження тиску в ротовій порожнині, і таким чином відбувається акт, подібний до кашлю, що спричиняє вилучення частинок із дихальних шляхів ссавців.
Дихання в повітрі. В атмосфері, як уже зазначалося, забезпечена висока і стала концентрація оксигену. Це дає можливість інтенсифікації процесів обміну речовин і перетворення енергії.
У тварин, які дихають повітрям, трапляються три головні типи дихальних органів: зябра, трахеї та легені.
Зябра. Ці органи не дуже добре пристосовані до дихання в повітрі. Тому вони використовуються небагатьма тваринами, зокрема тими, що відносно недавно перейшли до наземного способу життя. Типовим прикладом є краб пальмовий злодій Birgus latro, який має жорсткі зябра. Краб Cardiosoma може довго жити на суходолі та у воді. Ще один приклад тварин, які дихають зябрами на суходолі, — мокриці (Isopoda), що мешкають у вологих місцях і зябра яких вміщені в порожнину.
Деякі риби здатні дихати повітрям. Так, наприклад, звичайний вугор Anguilla може переповзати значні відстані суходолом і переселятися з однієї водойми в іншу, особливо вночі і по мокрій траві. Вугри на суходолі тримають свої зяброві порожнини наповненими повітрям, які оновлюють приблизно один раз за хвилину. У воді ці вугри роблять близько 20 дихальних рухів за хвилину. Для них характерна зміна значення зябер і шкіри під час дихання у воді та повітрі. Так, у воді близько 90 % усього оксигену надходить через зябра і 10 % — через шкіру. У повітрі через зябра надходить приблизно 30 % оксигену, а через шкіру — 70 %.
Прикладом риби, яка отримує оксиген із повітря в основному через шкіру, є муловий стрибун Periophthalmus. Дихає повітрям через рот електричний вугор Electrophorus electricus. Панцирна щука Lepisosteus osseus має зябра, а також використовує для дихання свій видозмінений плавниковий міхур, який періодично заповнює повітрям.
Особливу групу риб, які дихають повітрям, становлять двояко- дихаючі риби. Так, Protopterus і Leptodosiren мешкають у водоймах,
що можуть пересихати. Під час засухи вони перебувають у своїх капсулах сплячими в мулі, а за настання вологого періоду поновлюють нормальне життя. У Protopterus більша частина оксигену потрапляє через легені, менша — через зябра, у Leptodosiren майже весь оксиген надходить через легені і лише незначна частина — через зябра. Водночас у Neoceratodus головним органом дихання слугують зябра. Ця риба живе в річках і струмках, у засушливий період перебуває у сплячому стані. За відсутності води вона все ж таки заповнює легені повітрям, що істотно допомагає надходженню оксигену в організм, але використання О2 різко зменшується.
Трахеї. Цей тип органів дихання характерний для комах. Він становить собою систему трубочок, які підводять оксиген безпосередньо до тканини. Тому циркуляція крові для транспорту О2 непотрібна. Газообмін у трахеях здійснюється шляхом дифузії, у багатьох великих за розмірами й активних комах відбувається односпрямований рух повітря через трахеї. За цього способу газообмін набагато кращий, ніж почергове всмоктування й виштовхування повітря.
Трахеї сполучаються із зовнішнім середовищем у більшості комах отворами, які називають дихальцями. Вони можуть закриватися, що дозволяє контролювати газообмін між атмосферою та повітрям у трахейній системі. Трахеї розгалужуються і заходять у всі частини тіла. Найтонші з них — трахеоли — мають діаметр до 1 мкм. Трахейна система в деяких комах (зокрема, личинок поденок Ephemeroptera) є цілком замкнутою, без отворів на поверхні тіла. У цьому разі газообмін здійснюється дифузією через кутикулу комах. Ще одним різновидом трахейної системи є розміщення її в порожнині прямої кишки. Комаха вентилює трахеї, коли набирає воду в пряму кишку й виштовхує її назад. Таку систему мають личинки.
Вентиляція трахей забезпечується тонкостінними розширеннями (повітряними мішками), які сполучені з більшими трахеями й під час стискування виштовхують повітря.
У багатьох комах основні дихальні рухи виконує черевце. Під час польоту вентиляція посилюється рухами грудей. Дихальні рухи синхронні з відкриванням та закриванням дихалець і також контролюються нестачею О2 або надлишком СО2.
Деякі комахи, які мешкають у воді, зберегли повітряний тип дихання — мають трахеї, які сполучаються із зовнішнім середовищем дихальцями, що приводяться в контакт з атмосферним повітрям і розміщені на задньому кінці тіла. Дрібні комахи (наприклад, личинки комарів) мають закриту трахейну систему, і газообмін відбувається шляхом дифузії.
Багато водних комах несуть запас повітря на поверхні свого тіла, і дихальця відкриті в цей повітряний шар. Повітря утримують незмочувані поверхні тіла, які мають гідрофобні волоски. Дорослі жуки- плавунці Dytiscus мають великі повітряні мішки під крилами. Клопи Notonecta несуть запас повітря на черевній поверхні.
Ще один тип утримання повітря водяними комахами — гідрофобні волоски на поверхні тіла, яка має назву пластрон. Типовим представником таких комах є клоп Aphelocheirus, на поверхні пластрона якого міститься до 2 млн волосків.
У напівжорсткокрилих комах Microvelia buenoi і Anisops на черевці є великі клітини, заповнені гемоглобіном, що зв’язує оксиген. Цей оксиген використовується під час перебування комах під водою.
Легені. Ці дихальні органи розміщені в заглибленнях, випинаннях або в порожнинах тіла. Розрізняють дифузні та вентиляційні легені. Дифузним тип легенів характеризується тим, що обмін повітря з атмосферою відбувається тільки шляхом дифузії. Цей тип дихання притаманний для відносно невеликих за розмірами тварин — легеневих слимаків, скорпіонів, деяких раків тощо. Вентиляційні легені мають тільки хребетні тварини.
У земноводних легені — це своєрідний мішок, який розділений кількома гребінцями, що збільшують поверхню. Легені ссавців розділені на велику кількість маленьких мішечків — альвеол.
У легенях оксиген з альвеолярного повітря переходить у кров, а СО2 — з крові в легені шляхом дифузії через стінки кровоносних капілярів і альвеол.
Рух повітря дихальними шляхами здійснюється за рахунок ритмічних дихальних рухів. У їх здійсненні беруть участь діафрагма, зовнішні міжреберні м’язи, міжхрящові м’язи трахеї та бронхів. За глибокого вдиху одночасно скорочуються м’язи діафрагми, міжреберні м’язи, а також деякі (трапецієподібний, ромбоподібний, великий та малий грудні та ін.) м’язи грудей і плечового поясу, які піднімають ребра вище, ніж за спокійного вдиху. Після закінчення вдиху дихальні м’язи розслабляються і грудна клітка повертається до висхідного положення — відбувається видих. Спокійний видих здійснюється без участі м’язів. За глибокого видиху скорочуються внутрішні міжреберні м’язи та м’язи живота.
Органи дихання птахів відрізняються від органів дихання ссавців. Легені птахів з’єднуються з об’ємними тонкостінними повітряними мішками. У птахів розгалуження бронхіальної системи не закінчуються альвеолами, як у ссавців, а мають наскрізний прохід повітря, що дає змогу йому протікати безперервно через легені поблизу газообмінної поверхні.
Для забезпечення оксигеном ембріонів птахів відбувається дихання яєць. На тупому кінці яйця є повітряна камера, розміщена між внутрішньою і зовнішньою підшкаралупними оболонками. Цей повітряний простір поступово збільшується, коли яйце втрачає воду внаслідок випаровування. У твердій шкаралупі яйця з карбонату кальцію існують пори. Так, у середнього за розмірами курячого яйця — приблизно 10 000 пор.
Перед вилупленням пташеня проколює внутрішню підшкаралупну оболонку повітряної камери й починає дихати повітрям із цієї камери. Потім пташеня прокльовує шкаралупу яйця й дихає повітрям атмосфери.
Перенесення оксигену кров’ю. У багатьох безхребетних оксиген переноситься в гемолімфі або крові в розчиненому стані. Проте кількість оксигену, яка може переноситися таким способом, невелика. У ссавців вона становить приблизно 0,2 мл на 100 мл крові. Ви- сокоорганізовані тварини (усі без винятку хребетні) мають у крові речовини, здатні зворотно зв’язувати оксиген у кількості майже у 20 разів більше, ніж за його розчинення у крові.
Речовини, які слугують переносниками оксигену в крові, є ферум- або купрумвмісними білками (табл. 8.5).
У деяких тварин дихальні металовмісні білки розчинені в крові, а в інших (зокрема, хребетних) вони наявні в клітинах еритроцитів. У більшості ссавців цілі еритроцити не містять ядра й мають діаметр 5—10 мкм. У земноводних, плазунів, риб і птахів еритроцити мають ядро, їхні розміри в 100—200 разів більші, ніж у ссавців. Єдині хребетні, які не мають еритроцитів і гемоглобіну, — це личинки вугрів (лептоцефаліди) і деякі антарктичні риби родини Chaenichthyidae.
Роль шкіри в диханні. Газообмін через шкіру відіграє важливу роль у земноводних. Деякі дрібні саламандри Plethadontidae зовсім не мають легенів або зябер, і газообмін у них здійснюється в основному через шкіру, а також через невеликі ділянки слизової оболонки рота.
У жаб у зимовий період, коли використання оксигену невелике, шкіра поглинає більше його, ніж легені. У період їхньої активної діяльності поглинання оксигену за участі легенів значно вище, ніж через шкіру.
Шкіра плазунів, на відміну від шкіри земноводних, майже непроникна для оксигену. Тому вони дихають легенями. Водночас морські змії, зокрема Pelamis piaturus, поглинають до 33 % необхідного оксигену через шкіру.
У ссавців газообмін через шкіру надзвичайно малий. Поглинання оксигену майже не вимірюється сучасними методами, а виділяється тільки до 1 % СО2 від тієї кількості, яка виводиться легенями. У кажанів Eptesicus fuscus через шкіру крил за 18 °С виводиться 0,4 % СО2 від усієї кількості утвореної цієї сполуки. Однак при 27,5 °С виводиться вже 11,5 % СО2. Разом з тим у цих тварин поглинання оксигену через шкіру крил надзвичайно мале.
Таблиця 8.5
ДИХАЛЬНІ МЕТАЛОВМІСНІ БІЛКИ (за Шмідтом-Ніельсоном, 1982, зі змін.)
Металовмісні білки |
Характеристика металовмісних білків |
Деякі тварини, які містять металовмісні білки |
Гемоціанін |
Білок із вмістом купруму |
Молюски: хітони, головоногі, передньозяброві, черевоногі. Членистоногі: краби, омари Павукоподібні: Euscorpius |
Гемоеритрин |
Ферумвмісний білок непорфіринової структури, наявний у середині клітин |
Сипункуліди Поліхети: Magelona Приапуліди: Helicryptus, Priapuhis Плечоногі: Lingula |
Хлорокруорин |
Ферумвмісний білок, наявний у розчині |
Поліхети: Sabellidae, Seipulidae |
Гемоглобін |
Ферумвмісний білок, наявний у розчині або клітинах |
Найпростіші: Paramecium, Tetrahymena Плоскі черви: паразитичні сисуни Нематоди: Ascaris Кільчасті черви: Lumbricus, Tubifex, Arenicola, Spiroibis (деякі мають хлорокруарин, а деякі не мають металовмісних білків); Seipula (містять також хлорокруарин) Членистоногі: комахи (Chirono- mus, Gastrophilus); ракоподібні (Daphnia, Artemia) Молюски: Planorbis Голкошкірі: голотурії хребетні майже всі, за вийнятком лептоцефалід (личинок вугрів) і деяких антарктичних риб (Chaenichthys) |