БИОХИМИЯ И МОЛЕКУЛЯРНАЯ БИОЛОГИЯ - В. ЭЛЛИОТ - 2002
ГЛАВА 22. СИНТЕЗ, ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ И ДЕГРАДАЦИЯ БЕЛКОВ
В предыдущей главе мы познакомились с процессом транскрипции гена, который приводит к образованию мРНК. В этой главе мы обсудим процессы синтеза, адресной доставки и избирательной деградации белков. Вы узнаете, каким образом происходит процесс трансляции мРНК в белок. Большинство белков эукариотической клетки синтезируется в цитоплазме (за исключением белков митохондрий и хлоропластов), и для их адресной доставки в ядро, плазматическую мембрану, мембраны митохондрий и другие клеточные органеллы существуют специальные механизмы. Наконец, разные белки программируются для короткого или продолжительного времени жизни.
Основные принципы процесса биосинтеза белка
Сначала рассмотрим проблему в общих чертах. мРНК - достаточно длинная молекула, в ее состав входят 4 вида азотистых оснований. Белки синтезируются из 20 различных аминокислот, и последовательность оснований в мессенджере (мРНК) определяет последовательность аминокислот в белке. Код, состоящий из 1 азотистого основания, может кодировать только 4 аминокислоты; код, включающий 2 азотистых основания - 16 аминокислот, что по-прежнему явно недостаточно для 20. Таким образом, для кодирования 20 аминокислот необходимо минимум 3 азотистых основания. Каждая аминокислота в мРНК «записывается» в виде триплета азотистых оснований, называемых кодоном. 4 различных основания могут образовать 64 триплета, или кодона (4 • 4 • 4).
На первый взгляд, эволюция вполне могла ограничиться использованием 20 кодонов, игнорируя оставшиеся. Однако тогда в результате случайных мутаций неизбежно появлялись бы триплеты, не кодирующие аминокислоты. Это привело бы к бездействию генов, так как при встрече с триплетом мРНК, не кодирующим аминокислоту, синтез белка прекращается.
Эволюция выбрала альтернативную стратегию. Три кодона были зарезервированы в качестве стоп-сигналов, указывающих белоксинтезирующему аппарату, что синтез белка завершен. Эти 3 триплета (один из них - UАА) не кодируют аминокислот.
Если в результате мутации стоп-сигнал появится в кодирующей области мРНК, белок-продукт данного гена образовываться не будет. Если бы таких стоп-триплетов было больше (44), то мутации, приводящие к остановке синтеза белков, встречались бы гораздо чаще (впрочем, у бактерий стоп-мутации можно преодолевал путем мутаций супрессора, но на этом мы останавливаться не будем).
Все оставшиеся (числом 61) триплеты кодируют аминокислоты, значит, одну аминокислоту могут кодировать несколько различных триплетов. Это явление называется вырожденностью генетического кода. Как показано в табл. 22.1, только две аминокислоты - метионин и триптофан - имеют по одному кодону (для метионина - AUG).
Таблица 22.1. Генетический код
Основание 5′-конца |
Среднее |
основание |
Основание 3'-конца |
||
U |
C |
А |
G |
||
U |
UUU Phe UUC Phe UUA Leu UUG Leu |
UCU Ser UCC Ser UCA Ser UCG Ser |
UAUTyr UAC Tyr UAA Стоп* UAG Стоп* |
UGU Cys UGC Cys UGA Стоп* UGG Trp |
U С А G |
С |
CUU Phe CUC Phe CUA Leu CUG Leu |
CCU Pro CCC Pro CCA Pro CCG Pro |
CAU His CAC His CAA Gln CAG Gln |
CGU Arg CGC Arg CGA Arg CGG Arg |
U С А G |
А |
AUU Ile AUC Ile AUA Ile AUG Met+ |
ACUThr ACCThr АСА Thr ACG Thr |
AAU Asn AAC Asn AAA Lys AAG Lys |
AGU Ser AGC Ser AGA Arg AGG Arg |
U С А G |
G |
GUU Phe GUC Phe GUA Leu GUG Leu |
GCU Ala GCC Ala GCA Ala GCG Ala |
GAU Asp GAC Asp GAA Glu GAG Glu |
GGU Gly GGC Gly GGA Gly GGG Gly |
U С А G |
* Стоп-триплет не кодирует аминокислот + Триплет AUG является также кодоном, инициирующим трансляцию
Остальные аминокислоты имеют 2 и более кодонов; у лейцина их даже 6. Распределение кодонов не было случайным. В тех случаях, когда одну аминокислоту кодирует несколько кодонов, они похожи друг на друга. Например, кодоны изолейцина - AUU, AUC и AUA - различаются только по третьему основанию. Кроме того,
сходные триплеты кодируют аминокислоты, обладающие структурным сходством: например, алифатические аминокислоты изолейцин и лейцин (см. с. 38) имеют кодоны CUU (лейцин) и AUU (изолейцин). Благодаря этим особенностям генетического кода многие мутации, приводящие к изменению лишь одного основания, часто не влияют на структуру синтезируемого белка (изменение CUU на CUC не влияет на кодируемую аминокислоту - оба кодона соответствуют лейцину) или сопровождаются заменой на аминокислоты со сходной структурой (изменение CUU на AUU приводит к - замещению лейцина на изолейцин). Изолейцин и лейцин настолько схожи по размеру и гидрофобным свойствам, что такая замена может и не сказаться на- функции белка. Таким образом, организация генетического кода создает своеобразный генетический «буфер», при котором влияние многих точечных мутаций на синтезируемый белок сводится к минимуму.
Как транслируются кодоны
Синтез белка, называемый трансляцией, происходит на рибосомах. Их структуру и функции мы обсудим позже. Сейчас же мы познакомимся с трансляцией кодонов в аминокислоты.
В структуре аминокислоты и ее кодона нет физического или химического сходства, которое могло бы приводить к их непосредственной ассоциации. Поэтому было высказано предположение о существовании адапторных молекул, обеспечивающих соединение аминокислот с соответствующими кодонами. В дальнейшем оказалось, что в роли таких адапторов выступают небольшие молекулы РНК. Их назвали транспортными РНК, или тРНК.
Транспортные РНК, или тРНК
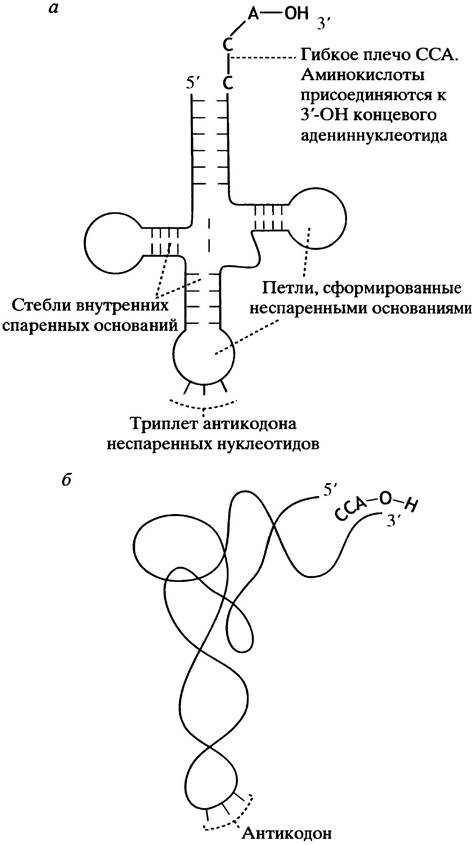
Это небольшие молекулы РНК. Схематически их структуру изображают в виде кленового листа (рис. 22.1, а). Благодаря спариванию оснований образуется двуспиральный стебель с петлей из неспаренных оснований. Наиболее важными частями считаются 3 неспаренных основания, которые формируют антикодон, и гибкое плечо - 3'-ССА, к которому присоединяется аминокислота.
В процессе биосинтеза 2 молекулы тРНК должны одновременно рядом поместиться на рибосоме, а их антикодоны - спариться с соседними кодонами на мРНК. В реальной жизни молекулы тРНК компактно свернуты (рис. 22.1, б). На рис. 22.2 представлена пространственная модель тРНК.
Рис. 22.1. Транспортная тРНК. а - Структура транспортной тРНК (кленовый лист) б - схематическое изображение свернутой молекулы тРНК
Структура молекул тРНК является как бы «вещью в себе». Хотя в тРНК есть модифицированные, необычные основания, основными частями являются антикодон и участок связывания аминокислоты. Антикодон - триплет оснований, комплементарный кодону (рис. 22.3). Таким образом, если кодоном на мРНК является UUU (кодирующий фенилаланин), то соответствующим антикодоном на молекуле тРНК будет ААА. Важно то, что данная молекула тРНК будет связывать только фенилаланин. Вскоре мы рассмотрим процесс присоединения аминокислоты. Как уже отмечалось, аминокислоты кодирует 61 кодон, поэтому можно предположить, что для трансляции необходимо столько же молекул различных тРНК. В действительности разновидностей тРНК меньше. Понятно, что их должно быть хотя бы по одной на 20 аминокислот. Некоторые молекулы тРНК могут узнавать несколько кодонов, которые, конечно, должны представлять одну и ту же аминокислоту. Это достигается за счет механизма неоднозначного соответствия.
Рис. 22.2. Структурная пространственная модель дрожжевой тРНКРhe. Цветом выделены конец ССА и петля антикодона

Механизм неоднозначного соответствия, или «качания»
До сих пор, рассматривая процессы репликации и транскрипции, мы видели незыблемость принципа Уотсона- Крика - комплементарного спаривания азотистых оснований. Поэтому небольшое отклонение от этого правила при спаривании оснований кодон-антикодон может внести некоторую неразбериху. Однако упомянутое отклонение относится только к первому основанию антикодона (за одним исключением, о котором позже). «Неправильное» спаривание этого основания есть результат гибкости близлежащей структуры тРНК (см. рис. 22.3), так что U в этом положении будет взаимодействовать с А или G на кодоне, а G- с С или U. Это и есть неоднозначное образование пар (см. рис. 20.17).
Рис. 22.3. Схематичное изображение спаривания оснований антикодона молекулы тРНК с кодоном мРНК. Для достижения антипараллельного спаривания молекула тРНК переворачивается. Поэтому структура тРНК, приведенная на рис. 22.1 (с 5'-концом слева), в спаренной форме находится в перевернутом виде (слева 3'-конец); мРНК всегда изображается 5'-концом слева
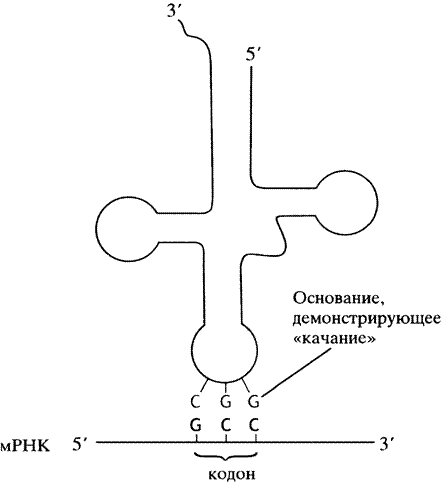
Понятие первое основание антикодона нуждается в разъяснении. Когда последовательность оснований антикодона приводится сама по себе, как и в случае любой другой нуклеотидной последовательности, она записывается в направлении 5' —> 3'. Однако при взаимодействии кодон-антикодон расположение триплетов антипараллельно, поэтому, если мы хотим показать тот же антикодон взаимодействующим с кодоном, последний всегда приводится в направлении 5' —> 3'; а «первое» основание антикодона в этом случае должно быть записано в противоположном направлении - справа. Вот почему на схемах молекула тРНК сама по себе приводится (на рис. 22.1) с 5'-концом, расположенным слева, но когда показывают ее спаривание с основаниями кодона, то 5'-конец расположен справа (на рис. 22.3). Таким образом, антикодон GGС будет взаимодействовать с кодоном как показано ниже:
![]()
Механизм «качания» допускает неправильное спаривание того же антикодона:

Поскольку и GCC, и GCU кодируют одну и ту же аминокислоту - аланин, такое спаривание не изменяет аминокислотной последовательности синтезируемого белка, но способствует трансляции одной тРНК нескольких кодонов. Это позволяет клетке синтезировать меньшее число молекул тРНК.
Эволюция позаботилась и о других путях, допускающих более гибкое взаимодействие кодон-антикодон без уменьшения точности трансляции. Один из них заключается в использовании в антикодоне нестандартного основания гипоксантина (см. рис. 18.7) для его спаривания с С, U или А в кодонах.
Как аминокислоты присоединяются к молекулам тРНК
Молекула тРНК присоединяет аминокислоту в соответствие с кодоном мРНК, комплементарному антикодону. Таким образом, молекула тРНК с антикодоном ААА «отвечает» только за фенилаланин, так как UUU - комплементарный ему кодон мРНК. Присоединение любой другой аминокислоты к этой тРНК привело бы к ошибке в структуре белковой молекулы: ведь в таком случае фенилаланин будет замещен другой аминокислотой. Специфичная для фенилаланина тРНК обозначается как тРНКPhе. тРНК остальных аминокислот записываются аналогичным образом, с использованием трехбуквенной аббревиатуры аминокислот (заметим, что тРНКРhе уточняет только вид тРНК, а не то, что Phe присоединен к ней: для этого используется обозначение Рhе-тРНКРhе). Присоединение аминокислот к тРНК катализируют ферменты аминоацил-тРНК-синтетазы. В каждой клетке должно быть по меньшей мере 20 различных видов этих ферментов, каждый из которых присоединяет определенную аминокислоту к соответствующей тРНК. Это означает, что каждый фермент узнает одну или несколько специфических тРНК и соответствующую аминокислоту, соединяя их вместе. Узнавание синтетазой соответствующей тРНК может происходить двумя путями: в одних случаях узнается антикодон, в других - несколько оснований в разных частях молекулы. Полная реакция, включающая АТР в качестве источника энергии, выглядит так:
Аминокислота + тРНК + АТР <-> Аминоацил-тРНК + РРi + АМР
Этот процесс иногда называют активацией аминокислот.
Неорганический пирофосфат гидролизуется до Рi, что обеспечивает протекание реакции слева направо. От правильного выбора синтетазами определенной аминокислоты зависит точность трансляции. После связывания тРНК со своей аминокислотой аминоацил-тРНК поступает в белоксинтезирующий аппарат, который уже не проверяет правильность выбора тРНК присоединенной аминокислоты. Поэтому очень важно, чтобы фермент, присоединяющий аминокислоты, при работе допускал мало ошибок. Активный центр фермента обычно высокоспецифичен относительно своего субстрата, однако пределы точности все же существуют. Фермент довольно легко отличает аминокислоты с сильно различающимися свойствами, но ему намного труднее распознать похожие аминокислоты, такие, например, как валин и изолейцин. Напомним их структуру.
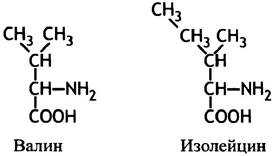
Различие в энергии связывания, обусловленное одной СН2-группой, не может обеспечить высокой селективности для изолейцина и валина. Без дополнительных мер предосторожности ошибка была бы слишком серьезной, и специфичная для лейцина аминоацил-тРНК- синтетаза присоединяла бы валин к тРНКIlе, приводя к неприемлемым ошибкам в трансляции мРНК. Впрочем, этого не происходит из-за наличия механизма коррекции, который можно описать в виде суммарной двухстадийной реакции.
Аминокислота + АТР Аминоацил-АМР + РРi (1)
Аминоацил-АМР + тРНК Аминоацил-тРНК + АМР (2)
Аминоацил-АМР не покидает фермента. Присоединение тРНК вызывает такое конформационное изменение молекулы фермента, при котором формируется
дополнительный каталитический участок, способный гидролизовать ошибочный аминоациладенилат. В случае ошибочного аденилирования изолейцин-специфичный фермент гидролизует валил-АМР, в то время как изолейцил-АМР (возможно, из-за гораздо больших размеров) в этот центр не входит. Такой механизм уменьшает ошибку присоединения изолейцина до 1 на 60 000 молекул. Другие аминоацилсинтетазы «вычитывают свою корректуру» аналогичным образом, гидролизуя ошибочные аминоацил-тРНК. Гидролитический центр соответствующей аминоацилсинтетазы узнает тРНК, несущие аминокислоты меньшего размера (по сравнению с правильным субстратом) или похожего размера, но содержащие другие боковые группы. Например, размеры треонина и валина схожи, однако гидролитический центр специфичного для валина фермента преимущественно связывает гидрофильную ОН-группу треонина, а не гидрофобную СН3-группу валина. Двойная избирательность - преимущественное связывание правильной аминокислоты с тРНК и преимущественный гидролиз аминоацил-тРНК, содержащих неправильную аминокисоту, - уменьшает ошибку до 1 молекулы на несколько тысяч. Не все аминоацил-тРНК-синтетазы имеют такие механизмы коррекции. Они нужны только для распознавания структурно похожих аминокислот. Случайная ошибка в биосинтезе белка не дает серьезных последствий, так как образуется множество молекул белка, и все они обязательно подвергаются деградации (дефектные молекулы, по-видимому, быстрее). Подобные ошибки не так критичны, как ошибки в синтезе ДНК; впрочем, высокая степень точности все-таки необходима.
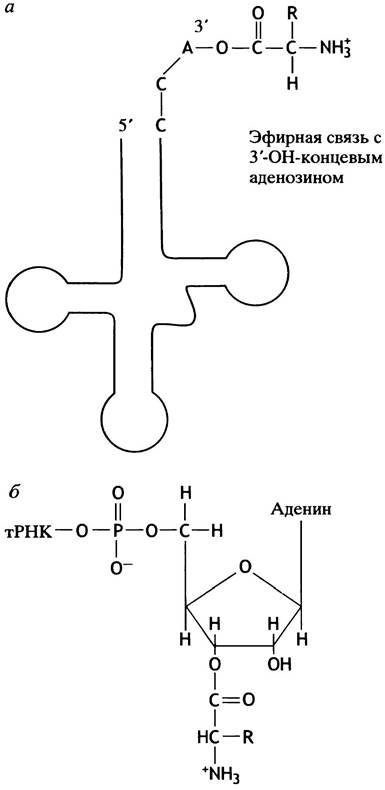
Молекула тРНК имеет концевую 3'-тринуклеотидную последовательность - ССА. Аминокислота соединяется эфирной связью со свободной 3'-ОН группой рибозы последнего нуклеотида (А) (рис. 22.4). Этот тринуклеотид образует гибкое плечо, которое может поместить аминоацильную группу на соответствующий реакционный центр рибосомы (см. ниже). Эфир, образованный аминоацильной группой, имеет тот же энергетический уровень, что и пептид. Таким образом, термодинамической проблемы переноса аминоацил-эфирной группы нa -NH2 другой аминоацильной группы при образовании пептидной связи не существует. Другими словами, энергия, необходимая для образования пептидной связи, обеспечивается аминоацил-тРНК-синтетазой, использующей АТР. Теперь познакомимся с организацией аминокислот на молекулах тРНК в полипептидную цепь.
Рис. 22.4. Присоединение аминокислоты к тРНК. а - Молекула тРНК с основаниями ССА на 3'-конце, к которой с помощью эфирной связи присоединена аминокислота; б - структура концевого нуклеотида с присоединенной аминокислотой. Аминоацильная группа в растворе быстро мигрирует между 2'- и 3'-ОН-группами цикла рибозы. В тексте мы рассматриваем ее в положении 3
Рибосомы
Рибосомы - маленькие частицы, присутствующие в больших количествах в клетках (за исключением высоко дифференцированных клеток, например, эритроцитов, которые белков не синтезируют). Свое название они получили от рибонуклеиновой кислоты (РНК), на долю которой приходится около 60% их сухого веса. Рибосома состоит из двух частиц: у Е. coli большая субчастица содержит 2 молекулы РНК, а малая субчастица - одну. Мы уже познакомились с мРНК и тРНК. Рибосомная РНК (рРНК) также имеет свои особенности. Из-за внутреннего спаривания оснований рРНК представляет собой сильно скрученные компактные структуры. Схема на рис. 22.5 поможет представить сложную форму одной из молекул РНК рибосомы Е. coli. Это - 16S рРНК малой субчастицы. Большая субчастица имеет две различные рРНК, оседающие при 23S и 5S. С рРНК ассоциированы многочисленные белки, в результате чего образуются плотные частицы; 34 белка входят в состав большой субчастицы и 21 - в состав малой.
Рис. 22.5. Структура 16S рРНК (схематично)
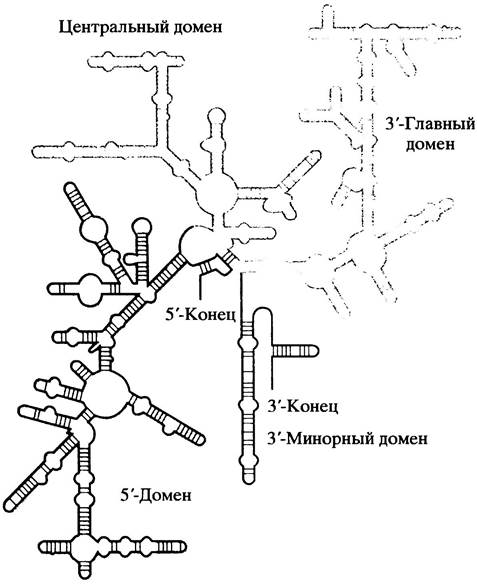
Когда речь идет об очень больших структурах, таких как рибосомы, их размеры оценивают по скорости седиментации (оседания) при ультрацентрифугировании, выраженной в единицах Сведберга (Svedberg), которые обозначаются S. Чем больше молекулы, тем быстрее они оседают. Размер рибосомы Е. coli - 70S, а ее субчастиц - 50S и 30S (суммарная величина не является результатом арифметического сложения субчастиц, так как величина S зависит не только от размера, но и от формы).
Общий принцип синтеза белка в сильно упрощенном виде выглядит так. Рибосома присоединяется около 5'-конца мРНК и затем движется вдоль мРНК по направлению к 3'-концу, соединяя аминоацильные группы нагруженных молекул тРНК (т. е. молекул тРНК с присоединенными молекулами аминокислот) в полипептидную цепь в соответствии с последовательностью кодонов. В конце мРНК рибосома встречает стоп-кодон: в этой точке белок высвобождается, рибосома отсоединяется и диссоциирует на субчастицы. Обратите внимание, что «особых» рибосом нет. В данной клетке любая рибосома может использовать любую мРНК, как плейер может использовать любую записанную ленту; ограничения существуют у рибосом митохондрий и хлоропластов, которые отличаются от цитоплазматических рибосом (см. с. 296).
Но это только принцип. Для понимания процесса нам необходимо остановиться на деталях. В ходе синтеза белковой молекулы можно выделить три стадии: инициацию, элонгацию и терминацию.
Инициация трансляции
Трансляция начинается со строго фиксированной точки в цепи мРНК, которая имеет 5'- и 3'- нетранслируемые области и расположенную между ними кодирующую область. Рибосома должна узнать первый триплет кодирующей последовательности и начать трансляцию с него. Необходима абсолютно точная инициация, поскольку правильная трансляция мРНК рибосомой зависит от точной установки рамки считывания. Предположим, что кодирующая область мРНК начинается с последовательности:
5' AUGUUUAAACCCCUG................. 3'.
Первые 5 аминокислот определены кодонами AUG, UUU, ААА и т. д. Нет никаких указаний на то, какие же основания составляют кодон; исключение составляют три первых основания кодирующей части мРНК, формирующие конститутивный кодон 1, следующая тройка - кодон 2 и т. д. Между ними нет ни запятых, ни точек. Следовательно, правильное считывание информации определяется лишь началом считывания. Предположим, произошла ошибка в одном основании, и мы начали считывание с основания 2. В этом случае кодоны читались бы так:
(A) UGU, UUA, AAC, ССС, UG ...
Иными словами, транслируемые кодоны, как и соответствующие им аминокислоты, были бы совершенно другими. Такая ошибка получила название сдвига рамки считывания.Мутации, вызванные делецией или добавлением одного или двух оснований в мРНК, вызывают сдвиг рамки считывания. Это приводит к тому, что аминокислотная последовательность пептида, синтезированного после сдвига рамки считывания, оказывается ошибочной, а образовавшийся продукт не способен
выполнять функции белка, отобранного миллионами лет эволюции и закодированного в данном гене. Общая схема синтеза белка и участвующих в нем компонентов в основном применима к про- и эукариотам, однако существуют значительные различия в реальных механизмах, поэтому мы рассмотрим их отдельно. Начнем с прокариот.
Инициация трансляции у Е. coli
Как уже отмечалось, перед стартовой точкой в мРНК есть последовательность, расположенная с 5'-конца цепи, которую определяет рибосома. Раз инициация не начинается с 5'-конца мРНК, то первый кодон должен быть каким-то образом идентифицирован.
Стартовой точкой является кодон AUG (реже GUG); однако, как ни странно, триплет AUG может располагаться в любой части мРНК, так как он кодирует аминокислоту метионин. Долгое время оставалось непонятным: каким же образом рибосомы инициируют трансляцию именно с первого AUG, а не с любого другого AUG в составе мРНК? Почему бы для инициации не использовать особый кодон? Оказывается, существует две различные тРНК, специфичные для метионина. Обе тРНК обладают одним и тем же кодоном, но одна тРНК используется только для инициации, а другая - только для добавления метионина в процессе элонгации.
Инициирующая трансляцию тРНК может спариваться с кодонами AUG или GUG за счет «качания» (см. с. 287). Обычно качание касается 5-основания антикодона. В используемой для инициации трансляции тРНК это 3'-концевое основание антикодона (рис. 22.6).
Рис. 22.6. Участок антикодона, демонстрирующий положение «качания». а - Обычные тРНК, участвующие в элонгации; б - инициаторная тРНКfМеt. Непостоянное основание (англ. wobble base) определяется близлежащей структурой тРНК. Значение тРНКfМеt объяснено в тексте
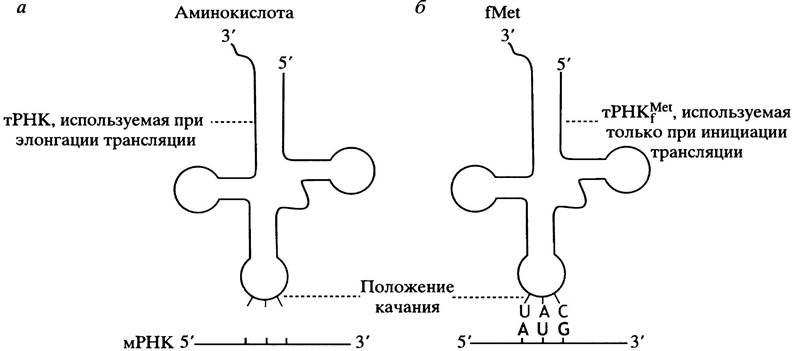
Что же определяет различные функции двух специфичных в отношение метионина тРНК? Инициаторная аминоацил-тРНК имеет структурные особенности, которые распознаются инициаторным белком, или фактором инициации IF2, доставляющим инициальную аминоацил-тРНК к формирующемуся инициаторному комплексу. Аминоацил-тРНК, участвующая в элонгации, распознает другой цитоплазматический фактор, который и доставляет их к рибосоме; он не связывается с инициаторной тРНК. У Е. coli метионин, присоединяемый к инициаторной тРНК, подвергается формилированию по NH2-группе. Этот процесс катализирует трансформилаза, использующая N10-формилтетрагидрофолат (см. с. 219) в качестве донора формильной группы, так что синтез белков прокариот, по непонятным пока причинам, начинается с N-формилметионина. Формильная группа (а часто и метионин) удаляются до завершения синтеза. Инициаторная тРНК обычно называется TPHKfMet («f» обозначает формил), а нагруженная аминокислотой тРНК - fMet-TPHKfMet (часто сокращается до fMet-TPHKf). Вышеизложенное проясняет, каким образом происходит правильное использование метионин-специфичных тРНК для инициации и элонгации: особые белковые факторы, участвующие в инициации и элонгации, узнают только Met-TPHKfMet и fMet-TPHKfMetсоответственно.
В цитоплазме существует фонд свободных 30 и 50S субчастиц рибосом, находящихся в равновесии с 70S рибосомами. Фактор инициации (IF3) связывается с 30S субчастицей и предупреждает на этой стадии ее преждевременную реассоциацию с 50S субчастицей (рис. 22.7, а). Кроме того, необходимы и другие факторы,
IF1 и IF2. Эти белковые факторы связываются с 30S субчастицами, участвуют в процессе инициации и освобождаются в цитоплазму, чтобы функционировать вновь в новом раунде инициации. Функция IF1 неизвестна, a IF2 необходим для связывания fMet-TPHKfMet.
Рис. 22.7. Влияние фактора инициации трансляции IF3 и последовательности Шайна-Дальгарно на субчастицы 70S рибосом. а - Диссоциация 70S рибосомы на 50S и 30S субчастицы. Фактор инициации IF3 предупреждает реассоциацию 50S и 30S субчастиц. При инициации трансляции IF3 должен быть высвобожден присоединения 50S;
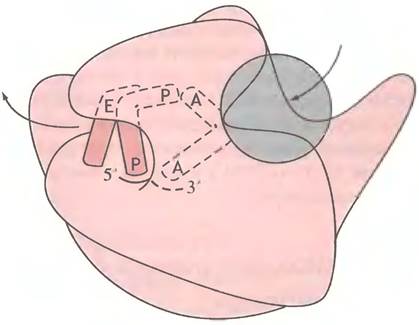
б - рисунок, показывающий роль последовательности Шайна- Дальгарно в расположении 30S рибосомы Е. соli на мРНК при инициации. Р - пептидильный участок, А - аминоацильный центр. Участки Р и А окончательно формируются только при присоединении 50S субчастицы
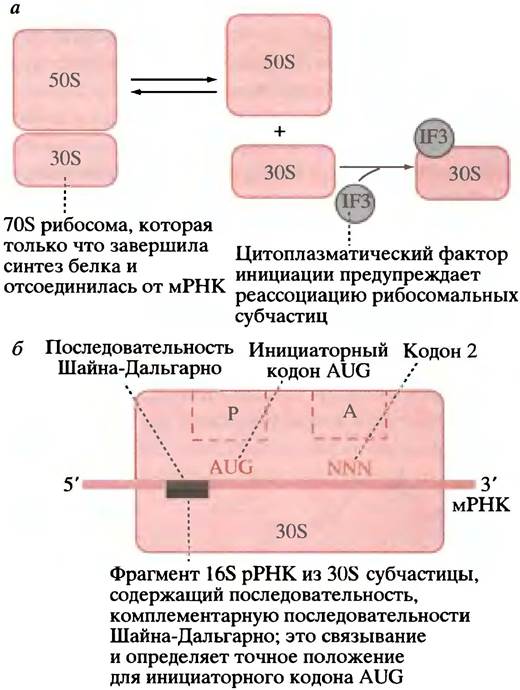
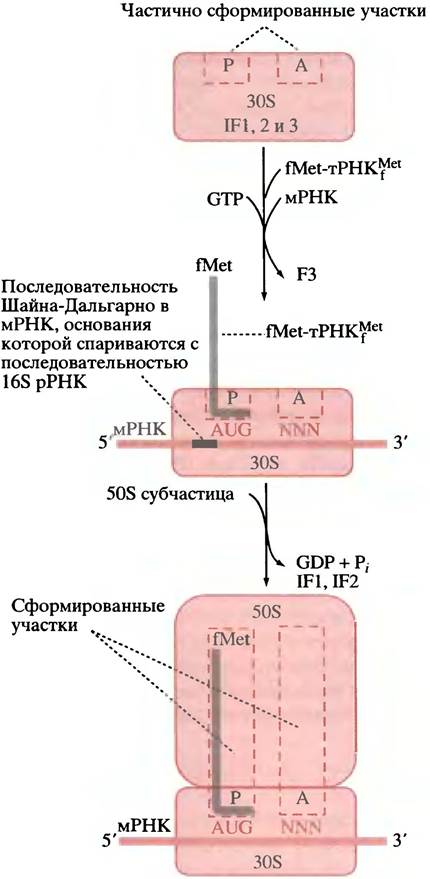
мРНК, fMet-TPHKfMet и GTP образуют комплекс с 30S субчастицей, a IF3 высвобождается в цитоплазму. Как показано на рис. 22.7, б, в мРНК существует последовательность оснований, известная как последовательность Шайна-Дальгарно, которая комплементарна участку 16S рРНК. При их связывании мРНК должным образом располагается на малой субчастице рибосомы, тРНК подходит к P-участку субчастицы, а ее антикодон спаривается с кодоном AUG (рис. 22.8).
После высвобождения IF3 этот комплекс соединяется с 50S субчастицей, что сопровождается гидролизом GTP и высвобождением GDP и Рр а также IF1 и IF2 (см. рис. 22.8). Теперь мы имеем полную 70S рибосому с мРНК, расположенной в P-участке с fMet-TPHKf (антикодон которой спарен с инициаторным кодоном AUG) и свободным A-участком, ожидающим доставки второй аминокислоты, соответствующей тРНК. Инициация завершена.
Рис. 22.8. Инициация трансляции у Е. coli Инициаторная тРНК - тРНКfМet показана серой линией; антикодон - короткой горизонтальной линией. Фактор IF2 доставляет fMet-TPHKfMetк 30S субчастице. NNN - любой кодон (N - любой нуклеотид). Рибосома имеет также центр выхода, не показанный на диаграмме
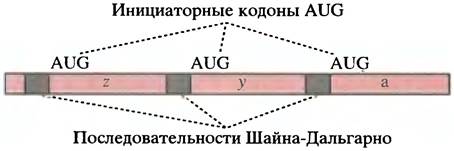
Некоторые бактериальные мРНК - полицистронны; например, lac мРНК. На одной молекуле мРНК находятся три области, кодирующие 3 различных белка. В этой ситуации каждая кодирующая область соседствует с последовательностю Шайна-Дальгарно, и в каждом случае инициация трансляции происходит независимо (рис. 22.9).
Рис. 22.9. Структура полицистронной мРНК прокариот. Кодирующие области lac мРНК обозначают z, y, и a (см. рис. 21.9)
Элонгация - следующий за инициацией этап трансляции
Цитоплазматические факторы элонгации
В цитоплазме есть два растворимых белковых фактора элонгации. Они являются G-белками, которые в комплексе с молекулой GTP могут связываться с рибосомами. Оба белка представляют собой латентные ОТРазы. На рибосоме они гидролизуют связанную молекулу GTP до GDP и Рi; освобождение последнего вызывает конформационные изменения в данных белках. В комплексе с GDP они отсоединяются от рибосомы. В цитоплазме GDP обменивается на GTP, и факторы готовы участвовать в следующем этапе элонгации.
Этими двумя факторами являются EF-Tu - нетермостабильный фактор элонгации (англ, elongation factor temperature unstable) и EF-G. Задача EF-Tu состоит в доставке аминоацил-тРНК к рибосоме. Как только аминоацильная группа добавится к растущему пептиду, EF-G способствует продвижению рибосомы по мРНК в направлении 5' —> 3' к следующему кодону.
Эти два фактора попеременно двигаются по рибосоме в связанном с GTP состоянии, выполняют свои задачи и отсоединяются в связанном с GDP состоянии, чтобы затем повторно участвовать в последующих раундах элонгации. А теперь мы можем перейти к детальному рассмотрению синтеза пептидной связи.
Механизм элонгации
Начнем с инициаторного комплекса, приведенного на рис. 22.10, а. В этом комплексе есть fMеt-тРНКfМеt в Р-участке и свободный А-участок. Только в процессе инициации Р-участок принимает тРНК, нагруженную аминокислотой - N-формилметионином; все последующие аминоацил-тРНК поступают в А-участок. Аминоацил-тРНК (отличные от инициаторных) образуют комплекс с фактором элонгации ЕF-Тu, связанным с молекулой GТР. Комплекс ЕF-Тu-GТР- аминоацил-тРНК связывается с рибосомой таким образом, что аминоацил- тРНК занимает А-участок, а ее антикодон располагается у кодона мРНК (рис. 22.10, б). Важно отметить, что EF-Tu-GTP не связывается с fMet-TPHKf(инициаторной тРНК), а взаимодействует только с аминоацил-тРНК, участвующими в элонгации. Таким образом, инициация и элонгация осуществляются раздельно. Концентрации EF-Tu в клетках Е. coli вполне достаточна для связывания всех аминоацил-тРНК. На рибосоме EF-Tu, обладающий ОТРазной активностью, гидролизует связанную молекулу GTP до GDP и Рi, высвобождая последний. Это служит сигналом для отделения EF-Tu от аминоацил-тРНК, по-видимому, вследствие аллостерических изменений, а EF-Tu-GDP покидает рибосому для регенерации в ходе обмена GDP-GTP в цитоплазме (рис. 22.10, в).
Рис. 22.10. Процесс элонгации. тРНК указаны в виде серых линий; АА - аминоацильная группа. Положение ЕF-Тu-GТР на тРНК и на рибосоме показано условно
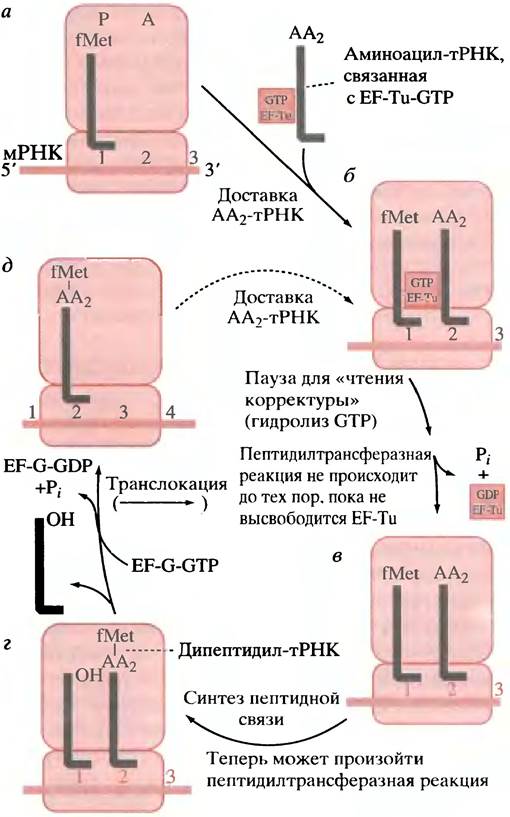
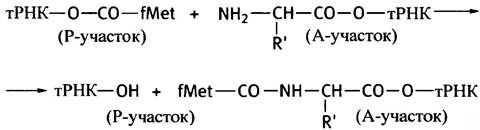
Аминоацильные группы на двух молекулах тРНК, расположенных в Р- и A-участках, находятся вблизи рибосомной пептидилтрансферазы: последняя катализирует перенос fMet от тРНК, расположенной в P-участке, на свободную аминогруппу аминоацил-тРНК в A-участке, образуя дипептид, присоединенный к тРНК (рис. 22.10, г). Мы описали пептидилтрансферазу как фермент, но есть данные, свидетельствующие о том, что эта активность сохраняется в рибосомах даже после экстракции 95% рибосомного белка. Это убедительный довод в пользу того, что данная активность является свойством рибосомной РНК.
Схема пептидилтрансферазной реакции выглядит так:
После синтеза первой пептидной связи A-участок заполнен пептидил-тРНК, а Р-участок содержит ненагруженную тРНК. Рибосома передвигается на один кодон вдоль мРНК: этот процесс называется транслокацией. Освободившаяся тРНК двигается к «выходу» — центру, расположенному на 50S субчастице, из которого она высвобождается для последующего использования (рис. 22.10, д). Для движения рибосомы по мРНК необходим фактор EF-G, который известен также как транслоказа. Для процесса транслокации необходим гидролиз GTP. Связывание EF-Tu-GTP и EF-G на рибосоме происходит таким образом, что в данный момент времени только один из факторов может быть связан с рибосомой. Таким образом, синтез пептида и транслокация происходят поочередно (друг за другом).
На рис. 22.10 не уточнен механизм, при участии которого пептидилтрансферазная реакция и транслокация проходят от стадии (в) до (г). Недавние исследования показали, что в ходе этого процесса каждая тРНК занимает на рибосоме два участка (рис. 22.11). После освобождения от пептидильной группы тРНК занимает участки Р и Е (от англ, exit - выход); антикодоновый конец молекулы по-прежнему находится в P-участке, но другой конец - уже в Е-участке. Аналогично этому тРНК, расположенная в A-участке (и несущая пептид), занимает А и Р-участки.
Обе модели основаны на наблюдении, что тРНК занимают участок Р/Е и А/Е на рибосоме. Согласно модели I, АА-тРНК раскачивается для того, чтобы сделать шаг; модель II предусматривает формирование на рибосоме гибридных участков. Предполагается, что синтез происходит таким образом, что пептид всегда остается в одном положении относительно большой субчастицы.
Для объяснения механизма этого процесса предложены две альтернативные модели. Модель I (см. рис. 22.11) предусматривает, что один конец аминоацил- тРНК раскачивается так, как показано на рисунке; происходит перенос пептида, а освободившаяся тРНК поворачивается одним из концов к Е-участку. Следующая затем транслокация вновь возвращает нас к ситуации, показанной на рис. 22.10, г, когда все готово для следующего раунда элонгации.
Рис. 22.11. Альтернативные модели транслокации и синтеза пептида на рибосоме. На этом рисунке для примера выбран синтез первой пептидной связи, так что серый кружок обозначает fMet, присоединенный к тРНК в Р-участке, который располагается у первого кодона мРНК; в конце этого раунда Р-участок располагается у второго кодона. Прямоугольники обозначают участки связывания, Е - центр выхода, Р - пептидильный участок, А - акцепторный участок. Е/Р и А/Р - гибридные центры, образованные движением большой субчастицы относительно малой
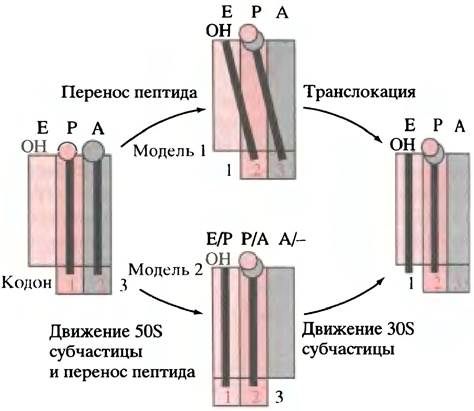
Согласно альтернативной модели II, движется большая субчастица рибосомы, а малая остается неподвижной. Это приводит к образованию гибридных центров связывания Р/Е и А/Р. После пептидилтрансферазной реакции гибридные участки оказываются занятыми (см. рис. 22.11). Транслокация малой субчастицы затем приводит к ситуации, предусмотренной моделью I, когда все готово к приему следующей аминокислоты.
Согласно обеим моделям, синтезирующийся пептид остается в фиксированном положении относительно большой субчастицы. Это позволяет преодолеть проблему физического передвижения тРНК, нагруженной достаточно большим пептидом. С помощью модели II можно также объяснить, почему рибосомы всегда состоят из двух субчастиц. Рибосома имеет очень сложную туннелеподобную структуру (рис. 22.12).
Рис. 22.12. Рибосома с вероятным расположением рибосом-связанных тРНК в А/А-, Р/Р- и Е-центрах (схематично). Серая область справа - примерный участок взаимодействия ЕF-Тu. На рисунке показана полярность фрагмента мРНК, содержащей кодоны в А- и Р-участках. Стрелки - вероятный путь тРНК на рибосоме
Итак, рибосома теперь находится в состоянии, показанном на рис. 22.10, д. Р-участок занят дипептидил-тРНК. А-участок, расположенный у третьего кодона, свободен. Элонгация полипептидной цепи включает повторные циклы одного и того же процесса. Амино- ацил-тРНК, образующая комплекс с ЕЕ-Ти-СТР, доставляется в А-участок, на нее переносится пептидильная группа с Р-участка; освободившаяся тРНК движется к «выходу» (Е-участок), пептидил-тРНК перемещается из А-участка в Р-участок, а рибосома перемещает один кодон - и весь процесс начинается вновь. Если синтезируется белок, насчитывающий 200 аминокислот, конечный раунд включает перенос пептида длиной 199 аминокислот на последнюю аминокислоту и ее тРНК, и это приводит к образованию комплекса белок-тРНК. Образно говоря, собака добавляется к хвосту, а не наоборот. Вначале синтезируется N-конец полипептидной цепи, а последняя присоединяемая аминокислота формирует С-конец.
Точность трансляции достигается взаимодействием кодон-антикодон, которое и определяет выбор соответствующей тРНК; однако пока неясно, каким образом происходит подобная селекция. Дело в том, что EF-Tu «не узнает», какая аминоацил-тРНК должна быть следующей, и похоже, что этот фактор доставляет их в А-участок беспорядочно. Существует гипотеза, согласно которой «неузнаваемый» комплекс амино- ацил-тРНК-EF-Tu-GTP диффундирует еще до реакции, поскольку он не будет удерживаться в А-центре также прочно, как это происходит при формировании водородных связей в ходе правильного взаимодействия кодона с антикодоном. Синтез пептида не происходит до тех пор, пока не будет высвобожден EF-Tu-GDP, а это может случиться только после гидролиза GTP. Предполагается, что небольшая задержка нужна для «вычитывания корректуры», т. е. для обеспечения эффективности процесса. Кроме того, рибосомы, как полагают, могут каким-то (пока невыясненным) образом участвовать в узнавании правильной аминоацил-тРНК. Но пока это только предположение, для объяснения которого еще не предложен механизм.
Терминация биосинтеза белка
На конце мРНК находится по крайней мере один из трех стоп-кодонов, называемых также терминаторными, для которых не существует tRNA: это триплеты - UAG, UAA и UGA. Когда рибосома достигает одного из стоп-кодонов, особый цитоплазматический фактор высвобождения белка (фактор терминации) способствует отделению готового полипептида от тРНК, вызывая изменения пептидилтрансферазы, в результате которых последняя гидролизует эфирную связь между СООН-группой белка, и ОН-группой 3'-концевого нуклеотида. Рибосома отсоединяется от мРНК и диссоциирует на субчастицы, готовые к следующей инициации. Известны два фактора, узнающих различные стоп-кодоны.
Что такое полисома?
Синтез одного белка Е. coli на рибосоме, присоединяющей около 15 аминокислот в секунду, занимает около 20 секунд. Однако, как только инициированная рибосома
продвинулась примерно на 30 кодонов, может произойти еще одна инициация трансляции, так что рибосомы одна за другой будут двигаться по одной мРНК, и каждая независимо друг от друга будет синтезировать белковую молекулу. Обычно на 1 молекулу мРНК приходится около 5 рибосом; впрочем, их количество изменяется в зависимости от длины мРНК. Такая структура из нескольких рибосом называется полисомой.
Чем отличается синтез белка у эукариот?
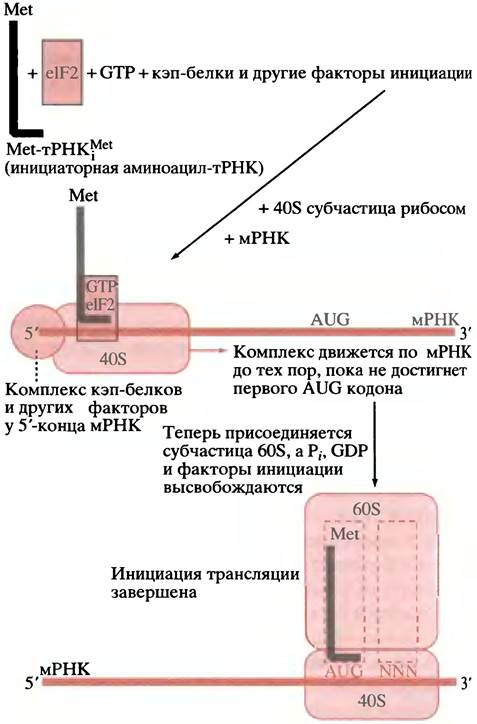
У эукариот процесс трансляции, в сущности, протекает так же, как и у прокариот, правда, с некоторыми отличиями. Рибосомы эукариот больше (80S, с субчастицами 40S и 60S) и содержат больше молекул рРНК и белка. Первой аминокислотой в синтезируемом белке всегда является метионин, однако метионил-тРНК, используемая для инициации, не формулируется (значение формулирования у прокариот непонятно). У эукариот, как и у прокариот, для инициации трансляции существует особая метионил-тРНК, которая отличается от тРНК, используемой в ходе элонгации. Это не означает, что все белки эукариот начинаются с метионина, так как часто эта аминокислота удаляется впоследствии из полипептидной цепи. Сборка 40S инициаторного комплекса аналогична образованию 30S инициаторного комплекса прокариот. Образовавшийся комплекс затем взаимодействует с большой 60S субчастицей рибосом, окончательно формируя инициаторный комплекс. Процесс биосинтеза белка также сопровождается гидролизом GTP. Однако у эукариот правильное расположение мРНК - строгое соответствие P-участка инициаторному кодону AUG - обеспечивает совершенно иной механизм. Напомним, что с 5'-конца мРНК эукариот кэпированы метилированным гуаниновым нуклеотидом (с. 270). В мРНК эукариот нет последовательности Шайна-Дальгарно; вместо нее к кэпу присоединяется группа белковых факторов, которая объединяется с 40S рибосомной субчастицей (рис. 22.13). Таким образом, с каждого 5'-конца мРНК на некотором расстоянии от кодона AUG мы имеем 40S преинициаторный комплекс. При помощи ATP-зависимого механизма комплекс 40S субчастицы движется вдоль мРНК до тех пор, пока не встретится первый триплет AUG: тогда происходит присоединение 60S субчастицы и завершение инициации. Этот процесс сопровождается гидролизом GTP. В отличие от мРНК прокариот, которые часто бывают полицистронными и содержат несколько последовательностей Шайна- Дальгарно, обеспечивающих инициацию трансляции каждого цистрона, мРНК эукариот - моноцистронны (кодируют один полипептид) и содержат только один инициаторный участок на молекулу. Функции прокариотических ЕF-Тu и ЕF-G у эукариот выполняют ЕF1α и ЕF2 соответственно.
Рис. 22.13. Упрощенное изображение инициации у эукариот. Помимо еlF2 в этом процессе участвует несколько факторов инициации. тРНКМеt - инициаторная РНК; еlF2 - фактор инициации эукариот, соответствующий фактору IF2 прокариот
Как происходит синтез белка в митохондриях?
Митохондрии содержат ДНК и имеют собственный белоксинтезирующий аппарат. Широко распространена гипотеза о том, что митохондрии будто бы произошли из прокариотических клеток, которые внедрились в клетки эукариот (с. 282).
Рибосомы митохондрий, подобно прокариотам, для инициации используют fMеt-тРНКfМеt. Для них характерны другие интересные особенности: несколько отличающийся генетический код и более простые взаимодействия кодона и антикодона, вследствие чего митохондрии млекопитающих могут обходиться 22 видами тРНК. Такие упрощения возможны, по-видимому, потому, что митохондрии синтезируют ограниченное количество различных белков. Митохондрия — не автономное образование. Большинство митохондриальных белков кодируются генами клеточного ядра и транспортируются в эти органеллы. Считается, что и хлоропласты также произошли из внедрившихся в эукариотические клетки фотосинтетических прокариот.
Влияние антибиотиков и токсинов на синтез белка
Антибиотики - химические «ракеты» микроорганизмов: последние выпускают их друг в друга в борьбе за выживание. Антибиотики атакуют наиболее важных участников клеточного метаболизма, некоторые из них задействованы и в процессе трансляции.
У прокариот стрептомицин нарушает инициацию, кирромицин предупреждает высвобождение ЕF-Тu, а эритромицин и хлорамфеникол ингибируют пептидил-трансферазу (последнее также справедливо и для митохондриальных рибосом). Фусидовая кислота тормозит транслокацию, блокируя высвобождение ЕF-G-GDР.
Кроме того, дифтерийный токсин ингибирует ЕЕ2-транслоказу эукариот (которая соответствует ЕF-G у бактерий). Рицин - токсин клещевины обыкновенной - это N-гликозидаза, удаляющая адениновое основание в одной из рибосомных РНК эукариот и инактивирующая большую субчастицу. Одна молекула рицина может разрушить клетку, содержащую десятки тысяч рибосом.
Как сворачивается полипептидная цепь, синтезированная на рибосоме?
Не так давно было принято считать, что после образования полипептидной цепи на рибосоме белок автоматически сворачивается в трехмерную структуру за счет взаимодействия аминокислотных остатков. Конечно, последнее очень важно, поскольку мы знаем, что структуру белка определяет только один тип информации - последовательность оснований в гене, которая в ходе трансляции преобразуется в аминокислотную последовательность полипептида. Более того, существует экспериментальное подтверждение этой концепции. Фермент рибонуклеазу денатурировали, обрабатывая мочевиной, нарушающей водородные связи, и восстанавливая дисульфидные связи. После удаления мочевины при диализе и реокислении часть фермента вновь правильно сворачивалась и даже проявляла каталитическую активность. Таким образом, аминокислотная последовательность белка, вне всякого сомнения, определяет его нативную конфигурацию. Однако в полипептидной цепи есть практически безграничные возможности ассоциации аминокислотных остатков друг с другом. Фрагмент из гидрофобных аминокислот в одной части цепи может - по мере синтеза белка - вступать во взаимодействие с другим гидрофобным фрагментом; однако этот контакт может быть совершенно «непригодным» и отсутствовать в нативном белке. Теоретически можно предположить, что полипептид «пробует» любой мыслимый вариант внутренней ассоциации, пока не достигнет свободной энергии нативного белка. Это беспорядочный метод фолдинга. Для такого процесса, как было подсчитано, необходимы многие миллионы лет, в то время как в клетке все происходит за считанные минуты.
После повторного сворачивания (рефолдинга) рибонуклеазы подобные эксперименты были проведены in vitro с другими небольшими белками, состоящими из одного домена. Попытки рефолдинга более крупных (особенно мультидоменных) белков были менее успешными. Поскольку большинство белков принадлежит к последней категории, очевидно, что сворачивание крупных молекул представляет серьезную проблему.
Механизм фолдинга белков in vivo, под которым мы понимаем пути приобретения вновь синтезированными пептидами правильной трехмерной конфигурации, окончательно не выяснен. Считается, что определенные фрагменты полипептида могут быстро принять вторичную структуру, и это каким-то образом облегчает правильный фолдинг всей молекулы. Последнее предположение лежит в основе концепции модульного фолдинга, при котором быстро образуются свернутые модули, способствующие фолдингу остальной молекулы. Таким образом, фолдинг белка, по-видимому, происходит в несколько этапов, однако детали этого процесса пока остаются невыясненными.
До сих пор мы обсуждали фолдинг белка как автономный процесс, осуществляющийся без посторонней помощи. Однако существуют два класса белков,
участвующих в фолдинге. К первому относятся традиционные ферменты, ко второму - молекулярные шапероны. В таком порядке мы их и рассмотрим.
Ферменты, участвующие в фолдинге белка
Первый фермент - протеин-дисульфидизомераза (ПДИ), которая «перемещает» S-S- связи в полипептидной цепи. Если образовалась ошибочная S-S-связь, то в силу своей ковалентной природы она не может разорваться спонтанно, и поэтому полипептид будет зафиксирован в неправильной конфигурации. Разрушая и вновь образуя S-S-связи между различными остатками цистеина, ПДИ способствует коррекции фолдинга. Высокие концентрации ПДИ обнаружены в эндоплазматическом ретикулуме, где происходит сворачивание белков, доставляемых для секреции, причем, многие из них имеют дисульфидные связи.
Другой фермент - пептидилпролин-изомераза (ППИ) - катализирует цис-, транс-изомеризацию пептидных связей пролина, что помогает белку принять правильную конфигурацию. Обнаружен целый класс совершенно удивительных белков, названных циклофилинами, которые обладают пролинизомеразной активностью. Они связывают антибиотик-иммунодепрессант циклоспорин (используемый при трансплантации органов). Связь между пролин-изомеразой и биологической ролью циклофилинов до сих пор непонятна.
Молекулярные шапероны и фолдинг
Шапероны (из англ.; букв. - пожилая дама, сопровождающая молодую девушку на балах) - семейство специализированных внутриклеточных белков, обеспечивающих быстрое нахождение правильной пространственной структуры. Они могут узнавать и связываться с частично свернутыми (или развернутыми) белками. При выходе из рибосомы в белке могут происходить «ошибочные» взаимодействия, например, между гидрофобными фрагментами; это будет препятствовать правильному сворачиванию молекулы. Связывание шаперонов с подобными фрагментами стабилизирует частично свернутую молекулу до того момента, пока не произойдет правильный фолдинг белка. Это означает, что шапероны должны «отойти» от полипептида: после диссоциации возникает благоприятная возможность завершения правильного фолдинга. Хотя детали этого механизма остаются неясными, очевидна важная роль шаперонов в фолдинге полипептидных цепей. О сложности системы свидетельствует и необходимость АТР для отделения шаперонов от полипептидной цепи. Обратите внимание, что шапероны имеют отношение к кинетике процесса фолдинга, а не к третичной структуре белка, определяемой аминокислотной последовательностью.
Болезни, вызываемые прионами, и фолдинг белка
Существует группа смертельно опасных неврологических болезней, поражающих человека и животных. Она включает болезнь Крюцфельда-Якоба и куру у человека. Последняя известна как «смеющаяся болезнь» из-за вызываемых ею гримас на лице. Предполагается, что в некоторых племенах Новой Гвинеи куру заражаются при каннибализме. У овец эту болезнь называют почесухой, потому что животные счесывают шерсть о столбы изгороди. У коров ее именуют губчатой энцефалопатией (англ. bovine spongiform encephalopathy; BSE), или болезнью коровьего бешенства. Эти болезни могут передаваться при употреблении в пищу инфицированной ткани. Реже встречаются наследственные заболевания. Раньше считали, что все перечисленные заболевания вызываются «медленными вирусами», потому что они заразны и развитие их происходит исподволь в течение ряда лет. Однако любые попытки обнаружить нуклеиновые кислоты в инфекционном материале, выделенном из мозга, оказались безуспешными, что позволило исключить возможность участия какого-либо инфекционного агента (например, вируса) в развитии заболевания. Впрочем, инфекционный агент, по-видимому, реплицируется.
Оказалось, что инфекция обусловлена устойчивой к протеазам формой нормального белка - прионовым белком, (англ. prion protein; РгР), обнаруженным в мозге. Вызывающий болезнь белок называется прионом по аббревиатуре от англ. proteinaceous infectious particle (белковая инфекционная частица).
Белок, заражающий мышей почесухой, называется PrPsc, а его нормальный «двойник» - РrРс («с» - от англ. constitutive, конститутивный). Оба представляют собой один и тот же полипептид и кодируются одним геном, однако их конформации различны: PrPsc характеризуется высоким содержанием β-слоев, которые практически отсутствуют у РrРс. В отличие от РrРс, белок PrPsc легко образует агрегаты, приводящие к образованию амилоидных бляшек, свойственных болезням, вызываемым прионами. Возникает вопрос: каким образом неправильно свернутый белок может стать инфекционным и воспроизводить себя? Ведь ни один из рассмотренных нами биохимических механизмов образования белка не позволяет ему управлять своей собственной репликацией. Тем не менее, существуют веские доказательства
того, что PrPsc каким-то образом переводит РrРс (нормальный белок) в аномальную форму. Это было подтверждено после совместной инкубации двух белков.
Модели механизма данного превращения подразумевают участие шаперона и источника энергии в раскручивании белка РrРс и его рефолдинг под влиянием молекулы PrPsc. Согласно альтернативной модели «образования ядра», молекулы РгРс захватываются агрегатом PrPsc, после чего и происходит конформационная реорганизация РrРс. Из-за ограниченных экспериментальных возможностей до сих пор нет доказательств приобретения белком in vitro инфекционных свойств в ходе описанных выше событий.
В отсутствие инфекции превращение РrРс в PrPsc происходит нечасто, поэтому спонтанные случаи заболевания встречаются чрезвычайно редко. Считается, будто мутации в гене, кодирующим нормальный РrР, могут увеличить вероятность этого посттрансляционного превращения, что приводит к появлению наследственных случаев болезни. Таким образом, однажды возникнув, PrPsc способен запустить свое дальнейшее аутокаталитическое образование.
Другие функции молекулярных шаперонов
Мы рассмотрели роль шаперонов в фолдинге белка, но это лишь часть выполняемых ими функций. Как уже отмечалось, шапероны способны присоединяться к несвернутым или частично свернутым белкам: полипептид, сошедший с рибосом, - один из них.
Однако белки могут частично денатурировать. Например, до определенной степени разворачиваться под действием различных факторов: тепла, чрезмерного окисления, ионизирующей радиации, ультрафиолетового света и т. д. Денатурацию белка также могут вызвать различные клеточные стрессы. Длительное время считалось, что тепловой шок индуцирует синтез особых белков, роль которых, как полагали, заключается в защите клетки. Если температуру Е. coli поднять до 42° С, произойдет вспышка синтеза белков теплового шока (с. 267). Оказалось, что это шапероны. Поэтому иногда их называют белками стресса.
Кроме участия в фолдинге вновь синтезированных белков, шапероны играют важную роль в разворачивании и стабилизации денатурированных белков, а также в мечении состарившихся белков с целью дальнейшей их деструкции лизосомами. Шапероны также необходимы для переноса белков через митохондриальные мембраны, для реорганизации субъединиц в комплексах и для сборки олигомерных белковых комплексов. Особенно удачное определение этого класса белков дали Хендрик и Хортл (см. раздел «Дополнительная литература»): «В настоящее время мы определяем молекулярный шаперон как белок, который связывается и стабилизирует неустойчивую форму другого белка. Контролируя связывание и высвобождение этого белкового субстрата, шаперон способствует образованию его правильной формы in vivo: будь это фолдинг, олигомерная сборка, транспорт в определенный компартмент клетки или контролируемое переключение активной/неактивной конформаций».
Последнее положение, касающееся переключения конформаций, относится к внутриклеточным рецепторам глюкокортикоидных гормонов (см. с. 346).
Для выполнения столь разнообразных функций нужны различные шапероны, и в настоящее время уже известны многие из них.
Как синтезированные белки доставляются по назначению?
В клетке существует только один тип рибосом, и все они локализованы в цитоплазме клетки (за исключением рибосом прокариотического типа, локализованных в митохондриях и хлоропластах). Какой белок будет в данный момент времени синтезироваться на рибосоме, зависит только от мРНК, которая в данный момент времени транслируется. У вновь синтезированных белков различные конечные пункты назначения. Многие из них являются цитоплазматическими, после синтеза высвобождаются из рибосом и остаются в цитоплазме; вместе с тем многие белки локализованы в мембране клетки. Как они туда попадают? Рассмотрим клетку печени: она продуцирует белки плазмы крови. Это крупные белки, а клеточные мембраны устроены так, что не допускают «утечку» белков. Каким же образом происходит избирательное высвобождение именно белков крови? Аналогичный вопрос возникает и при высвобождении любых внеклеточных белков, например, пищеварительных ферментов или инсулина из поджелудочной железы. Есть и другие проблемы. Большинство митохондриальных белков синтезируется в цитоплазме. Каким образом происходит их избирательная транспортировка в митохондрии? Лизосомы и пероксисомы (см. главу 16) - окруженные мембраной пузырьки, наполненные ферментами; но они не могут синтезировать белки. Каким образом последние транспортируются в эти пузырьки?
Сейчас мы познакомимся с различными типами перемещения (транслокации) белков. Начнем с тех, которые сначала транспортируются в эндоплазматический ретикулум (ЭР).
Что такое эндоплазматический ретикулум (ЭР)?
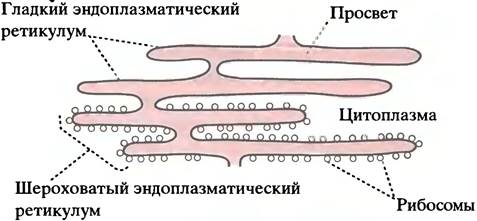
Если посмотреть на срез печени или поджелудочной железы в электронный микроскоп, внутри клетки можно увидеть большое количество мембран, ограничивающих огромное количество сплющенных полостей. Эта сложная сеть взаимосвязанных полостей называется эндоплазматическим ретикулумом (ЭР). Цитоплазма находится снаружи ЭР, а просвет ЭР представляет собой отдельный компартмент, ограниченный мембраной ЭР (см. электронную микрофотографию на рис. 3.15). Часть ЭР усеяна рибосомами, обусловливающими шероховатость ЭР в электронном микроскопе: это шероховатый эндоплазматический ретикулум. У части ЭР рибосомы отсутствуют, поэтому такой ЭР называют гладким эндоплазматическим ретикулумом (рис. 22.14). Белки, предназначенные, например, для ядра или митохондрий, сначала транспортируются в просвет шероховатого эндоплазматического ретикулума; это подводит нас к следующему вопросу.
Рис. 22.14. Строение эндоплазматического ретикулума
Как белки проходят через мембрану ЭР?
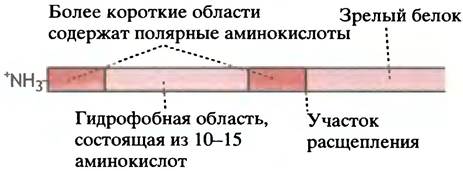
Белок, который должен проникнуть через плазматическую мембрану, внутрь лизосом или в просвет ЭР, имеет на N-конце лидерную последовательность, обычно состоящую из 25 ± 11 аминокислот. Лидерная аминокислотная последовательность определяет общее структурное свойство многих белков, транспортируемых в различные компартменты (рис. 22.15). Зрелый белок высвобождается после протеолитического расщепления в участке со специфической последовательностью.
Рис. 22.15. Присоединение типичной лидерной последовательности к N-концу белка, транспортируемого через мембрану ЭР. Подобные лидерные последовательности образованы определенным типом аминокислот, но не обладают специфической аминокислотной последовательностью; кислые аминокислоты в ней отсутствуют
Обратите внимание, что в цитоплазме свободная рибосома сначала синтезирует лидерную последовательность белка, которая сразу же распознается сигнал-узнающей частицей (англ. signal recognition particle; SRP). Последняя представляет собой цитоплазматический комплекс РНК-белок. Эта частица связывается с комплексом рибосома-синтезирующийся пептид, препятствуя дальнейшей элонгации полипептидной цепи. Механизм, обеспечивающий перенос «арестованной» рибосомой растущего пептида через мембрану ЭР, довольно сложен и до конца не изучен; впрочем, ряд важных моментов уже известен (рис. 22.16).
Рис. 22.16. Перенос полипептида через мембрану ЭР при участии мембранного транслокона
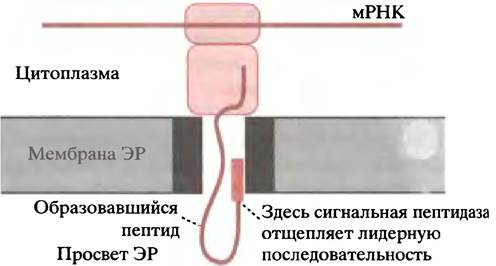
В мембране ЭР есть рецепторы SRP, или так называемые докинг-белки, к которым присоединяется цитоплазматический комплекс рибосома-SRP. В результате серии последовательных этапов, в ходе которых происходит гидролиз одной молекулы GTP до GDP, рибосома присоединяется к мембранному белку, который образует полипептид-транслоцирующую пору, иногда называемую транслоконом (см. рис. 22.16), а SRР освобождается в цитоплазму для повторного использования. Растущий полипептид переносится через белковую пору в мембране в виде петли с сигнальным пептидом, который прикрепляется к поре (см. рис. 22.16). Расположенная рядом с порой сигнальная пептидаза отщепляет лидерную последовательность, а полипептид поступает в просвет ЭР.
Пока неизвестно, что же управляет транслокацией растущего пептида. В фолдинге полипептида, по-видимому, участвуют находящиеся в просвете ЭР шапероны.
Гликозилирование белков в просвете ЭР
В главе 3 (см. с. 62) мы упоминали, что некоторые белки (особенно мембранные и секреторные) соединены с олигосахаридами. Местами присоединения являются либо NН2-боковые группы аспарагина (N-гликозилирование) или ОН-группы остатков серина и треонина (О-гликозилирование). Присоединение осуществляется в несколько этапов. Внутри ЭР происходит N-гликозилирование. Первый этап этого процесса особенно интересен тем, что «ядро» олигосахарида, состоящее из 14 мономеров сахара, образуется в цитоплазме и транспортируется через мембрану будучи присоединенным к длинной гидрофобной цепи. Последняя включает более 100 атомов углерода и называется долихолфосфат. Расположенный на внутренней стороне мембраны ЭР фермент трансфераза переносит олигосахаридную группу на образующийся пептид, как только тот появляется в просвете ЭР. О-гликозилирование происходит в аппарате Гольджи (см. следующий раздел).
Что происходит с полипептидом, перенесенным в просвет шероховатого ЭР?
ЭР представляет собой «мешок», из которого нет выхода. В просвете шероховатого ЭР белок окружается мембраной, и образующиеся пузырьки отделяются от не содержащих рибосомы областей ЭР, доставляя содержимое просвета ЭР в другую органеллу - комплекс Гольджи (аппарат Гольджи) путем слияния с ее мембраной.
Структура аппарата Гольджи проста: около полудюжины вытянутых мембранных мешочков, не имеющих ни входа, ни выхода. При помощи рассмотренных выше транспортных пузырьков он получает белки из ЭР и посылает их по назначению, отпочковывая пузырьки, как показано на рис. 22.17. Электронная микрофотография аппарата Гольджи приведена на рис. 3.15.
Рис. 22.17. Схематическое представление центральной роли аппарата Гольджи в посттрансляционной сортировке и мечении белков. Вновь синтезированные мембранные липиды метятся для доставки транспортных пузырьков точно по назначению. Образование пузырьков для транспорта между цистернами аппарата Гольджи происходит при участии удивительного механизма: комплекс GTP-белок взаимодействуют с мембраной, что приводит к связыванию «покровных» белков. Отпочковывание пузырьков от участков связывания ведет к образованию покрытых везикул. «Раскрываются» они при контакте с мембраной- мишенью в результате гидролиза GTP до GDP; «раскрытие» приводит к слиянию везикулы и мембраны-мишени. Молекулы-мишени на везикуле направляют последнюю точно по назначению. Механизм «укрывания» пузырьков, по-видимому, широко распространен
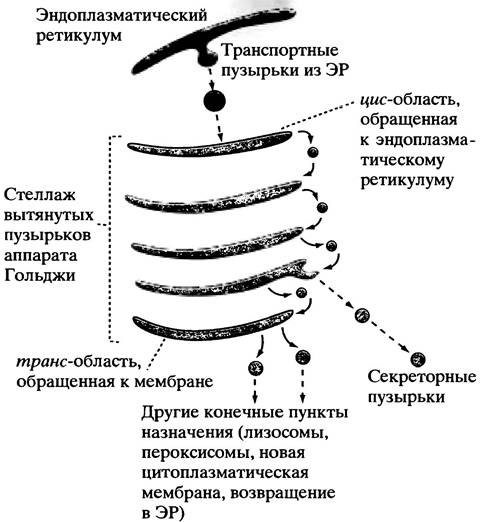
Не все белки, поступающие в аппарат Гольджи, предназначены для секреции. Конечным пунктом назначения одних являются лизосомы или пероксисомы. Другие, например, протеин-дисульфид-изомераза (с. 298) или ферменты гликозилирования, должны вернуться в шероховатый ЭР, потому что они оказались «унесенными» в аппарат Гольджи случайно, вместе с остальными белками. Аппарат Гольджи сортирует эти белки, «упаковывает» их в пузырьки, которые и доставляют содержимое в нужное место. Поэтому аппарат Гольджи, пожалуй, наиболее удивительное функциональное «подразделение» клетки, способное к сортировке белков по их конечному назначению и отправлению последних в «посылках» по точно указанным адресам.
Предназначенные для секреции белки аппарат Гольджи упаковывает в пузырьки, мигрирующие по направлению к плазматической мембране. Есть два типа секреции. Некоторые белки высвобождаются постоянно по мере образования: к ним относятся белки крови, выделяемые из клеток печени без дополнительных сигналов. Для этого пузырьки сливаются с плазматической мембраной клетки по мере поступления, а освобождение их содержимого осуществляется посредством экзоцитоза. Высвобождение пищеварительных ферментов, например, из поджелудочной железы, необходимо только при поступлении пищи в кишечник. В этом случае пузырьки имеют больший размер. Они хранят ферменты до тех пор, пока нейрональный или гормональный стимул не вызовет их массового освобождения в ходе экзоцитоза секреторных везикул (сигнал и механизм этого процесса описаны на с. 358). Отпочковывание транспортных пузырьков из мешочков аппарата Гольджи происходит после того, как мембраны будут окружены покровным белком в комплексе с G-белком и присоединенным к нему GТР. Гидролиз GТР служит сигналом для раскрытия и слияния везикул с мембраной-мишенью. В процессе узнавания важную роль играют особые белки, расположенные на везикулах и мембранах-мишенях.
Что такое «адресные сигналы» на белках, участвующих в сортировке в аппарате Гольджи?
Как описано ниже, в случае ферментов, направляемых для включения в лизосомы, сигнал несет углеводная часть гликопротеина. Однако в большинстве случаев механизм сортировки должен включать рецепторы, узнающие определенные структурные свойства или специфические последовательности белков, предназначенных для сортировки. Они еще окончательно не установлены, но одна категория рецепторов уже известна. У тех белков (например, у протеин-дисульфид-изомеразы), которые должны быть возвращены в ЭР, «меткой» служит последовательность, состоящая из четырех аминокислот: Lуs-Аsр-Glu-Lеu. В системе однобуквенного обозначения аминокислот (табл. 22.2) эта последовательность записывается как КDЕL.
Таблица 22.2. Однобуквенные символы аминокислот
Аланин |
А |
Лейцин |
L |
Аргинин |
R |
Лизин |
К |
Аспарагин |
N |
Метионин |
М |
Аспарагиновая кислота |
D |
Фенилаланин |
F |
Цистеин |
С |
Пролин |
Р |
Глутамин |
Q |
Серин |
S |
Глутаминовая кислота |
Е |
Треонин |
Т |
Глицин |
G |
Триптофан |
W |
Гистидин |
Н |
Тирозин |
Y |
Изолейцин |
I |
Валин |
V |
Упаковка лизосомных белков
До сих пор мы знакомились с белками, предназначенными для секреции. Внутриклеточный путь секреторных белков похож на путь лизосомальных белков. Как описано в главе 16, лизосомы - небольшие окруженные мембраной внутриклеточные органеллы, содержащие группу высокоактивных гидролитических ферментов. Ферменты образуются на ЭР тем же способом, что и секреторные белки, и в просвете ЭР они подвергаются N-гликозилированию.
В аппарате Гольджи ферментная система узнает лизосомные ферменты и фосфорилирует маннозу олигосахаридной части последних. В мембране аппарата Гольджи есть внутренние рецепторы, к которым прикрепляется фосфорилированная манноза; после этого происходит отпочковывание пузырьков, содержащих лизосомные ферменты. Пузырьки сливаются с другими «сортировочными пузырьками», внутри которых поддерживается низкое значение pH (за счет протонных насосов в мембранах). Кислый pH пузырьков способствует диссоциации рецепторов маннозофосфата от гликопротеинов, и они возвращаются (путем отпочковывания пузырьков) назад, в аппарат Гольджи; в это время ферменты доставляются к лизосоме. При генетическом заболевании, называемом болезнью I-клеток («I» обозначает включение мукополисахаридов), фосфорилирующая маннозу система дефектна, что приводит к нарушению поступления группы лизосомных ферментов в лизосомы. Последнее, в свою очередь, приводит к нарушению деградации мукополисахаридов, накопление которых в лизосомах сопровождается развитием серьезной клинической симптоматики.
Когда было обнаружено, что фосфорилирование остатков маннозы является своеобразным «ярлыком» лизосомного фермента, появилось предположение, что углеводное «мечение» должно быть широко распространено. Однако в большинстве случаев сортирующий механизм аппарата Гольджи узнает определенные структурные особенности белков.
Каким образом метятся интегральные белки цитоплазматических мембран?
Интегральные белки мембран также синтезируются на шероховатом ЭР, включаются в мембрану ЭР, транспортируются в аппарат Гольджи в составе везикул и оттуда (также в составе пузырьков) направляются к плазматическим мембранам. Синтез мембранных липидов также происходит в ЭР, и новые мембраны образуются в этой органелле.
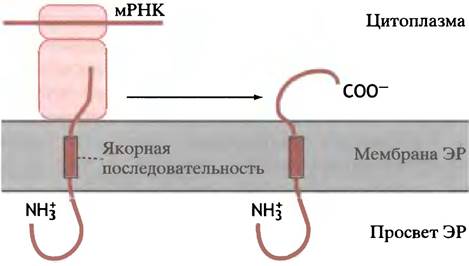
Интересен, конечно, вопрос: почему в случае секреторного белка полипептид идет прямо через мембрану шероховатого ЭР, а интегральный белок заякоривается в ней? Вся необходимая информация, которая определяет, будет или нет (и каким образом) данный пептид интегрироваться в структуру мембраны, заложена в аминокислотной последовательности белка.
Наиболее проста ситуация, когда в полипептиде есть якорная последовательность (называемая также останавливающей перенос) (рис. 22.18). Она фиксируется в гидрофобной части бислоя и останавливает последующий перенос. Затем рибосома завершает синтез С-концевой части полипептида, приводя к образованию белка, N-конец которого находится снаружи ЭР, а С-конец выступает в просвет ЭР. Когда участок ЭР становится фрагментом клеточной мембраны, этот белок имеет ту же ориентацию, а N-конец выступает снаружи клетки. Мембранные белки могут также иметь и противоположную ориентацию, «прошивая» мембрану несколько раз. Механизм встраивания таких белков предусматривает участие несколько лидерных и останавливающих перенос сигналов.
Рис. 22.18. Иллюстрация роли якорных сигналов во встраивании интегральных мембранных белков в мембрану ЭР. N-Конец полипептида ориентирован наружу, а С-конец - внутрь. Мембранные белки, участвующие в транслокации, не приведены. Другая организация якорных сигналов может способствовать расположению белков в противоположной ориентации
Является ли перенос всех белков ко-трансляционным?
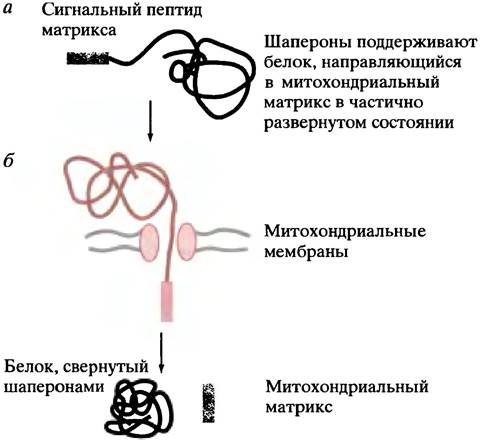
До сих пор мы рассматривали перенос (транслокацию) белков в ходе их синтеза через или в мембраны. Однако в клетках эукариот существует и другой тип транспорта белка - в митохондрии или хлоропласты у растительных клеток. Как уже отмечалось, у митохондрий есть хромосома и собственная белоксинтезирующая машина, но они ответственны лишь за синтез небольшой части белков этих органелл. Несколько сотен различных белков или субъединиц транспортируются в митохондрии, составляя более 95% присутствующих там белков. Они синтезируются в цитоплазме. По аналогии с ЭР можно было бы предположить, что синтез белка происходит на связанных с митохондриями рибосомах, однако дело обстоит не так. Митохондриальные белки синтезируются на свободных рибосомах. Кодируемые в ядре белки митохондрий (т. е. белки, кодируемые генами клеточного ядра) синтезируются и освобождаются в цитоплазму в виде пре-белков, после чего к ним присоединяются шапероны, поддерживая их в развернутой форме. Высвободившиеся (с рибосомы) белки пересекают мембрану в вытянутой (несвернутой) форме, и этот процесс управляется мембранным потенциалом. После того как белок пересечет мембрану, другие шапероны внутри митохондриального матрикса осуществляют его сворачивание.
Лидерная последовательность митохондриального белка насчитывает примерно 12-70 аминокислот. Сами последовательности варьируют от одного пре-белка к другому, но общим структурным свойством является амфипатическая α-спираль, в которой одна сторона заряжена положительно, а другая в значительной степени гидрофобна. Она присоединяется к рецепторам внешней митохондриальной мембраны.
В простейшем случае N-концевая лидерная последовательность направляет белок в матрикс митохондрий через пору, расположенную на внешней и внутренней мембранах в точках контакта. Затем лидерная последовательность отщепляется (рис. 22.19). В том случае, когда пункт назначения находится в межмембранном пространстве, пре-белок имеет две лидерных последовательности. Как уже было описано, первая направляет его в матрикс. Но ее удаление приводит к экспонированию второй, которая направляет полипептид из матрикса через внутреннюю мембрану в межмембранное пространство. Здесь происходит удаление второй лидерной последовательности.
Рис. 22.19 Перенос митохондриальных пре-белков. а - Схема строения митохондриального пре-белка, синтезированного в цитоплазме; конечный пункт назначения - матрикс; б - полипептид пересекает митохондриальную мембану через белковую пору, а лидерная последовательность удаляется сигнальной пептидазой матрикса.
Начальное прикрепление полипептида к митохондрии происходит за счет рецепторного белка в мембране (не показано). Если пунктом назначения является межмембранное пространство, вторая лидерная последовательность управляет движением белка из матрикса через внутреннюю мембрану митохондрий
Ядро клетки окружено двойной ядерной мембраной. Белки, синтезируемые в цитоплазме, переносятся в ядро через специальные поры. Во многих случаях для прохождения белков через эти поры необходима сигнальная последовательность, содержащая короткий аминокислотный фрагмент; для транспорта небольших белков-гистонов он не требуется. В случае доставляемых в ядро факторов транскрипции сигнальная последовательность находится внутри полипептидной цепи.
Не нужно путать пре-белок с про-белком! Инсулин секретируется в просвет ЭР в виде одиночной полипептидной цепи - проинсулина. В ходе удаления среднего фрагмента проинсулина он превращается в 2 полипептида, соединенных дисульфидной связью. мРНК инсулина кодирует лидерную последовательность проинсулина, что привело к появлению термина пре-проинсулин, хотя - поскольку пре-последовательность (лидер) удаляется при пересечении мембраны ЭР - пре-проинсулина как такового в клетке не существует.
Деградация белков
Мы познакомились с синтезом белка и его транслокацией. Теперь рассмотрим деградацию белка.
Важно отметить следующее: 1) деградация белка происходит во всех клетках, поскольку старые молекулы белка замещаются новыми; 2) деструкция белковых молекул высокоселективна. Некоторые белки имеют время полужизни более 20 часов (белки печени - несколько дней), другие - 10 и даже 2 минуты.
Есть несколько причин, обусловливающих деградацию белков. Поскольку абсолютная точность в трансляции, по-видимому, невозможна, неизбежно образуются белковые молекулы, содержащие ошибочную аминокислоту, что может привести к неправильному фолдингу молекулы. Белки, пострадавшие от химического повреждения, например, окисления, также должны быть уничтожены.
Однако «удалением отходов» эта тема не исчерпывается. Некоторые белки, похоже, намеренно созданы для быстрого разложения: об этом свидетельствует
присутствие определенных аминокислотных последовательностей. Почему клетка должна заботиться о быстрой деградации очень хороших ферментов? Во многих случаях в интересах метаболического контроля. Если фермент разрушается быстро, то его уровень можно тонко и постоянно контролировать скоростью его синтеза. Такие ферменты характеризуются относительно коротким временем жизни. Структурные белки и гемоглобин, наоборот, относятся к «долгожителям».
В одном из механизмов селективной деструкции белков участвует убиквитин - небольшой белок, найденный у всех эукариот, но не бактерий; так что его название (от англ. ubiquitous - вездесущий) явно преувеличено. В ходе ATP-зависимой реакции его концевая карбоксильная группа связывается с ε-аминогруппой боковой цепи остатков лизина белка-мишени, и последний таким образом оказывается помеченным для деструкции (рис. 22.20). Выбор убиквитином белков, предназначенных для разрушения, по-видимому, определяется их N-концевой аминокислотой. Шапероны тоже могут участвовать в мечении белков, подлежащих деструкции.
Рис. 22.20. Убиквитин-зависимое мечение белков, подлежащих деградации. Механизм выбора белков для деструкции неизвестен, хотя отмечено, что концевая аминокислота влияет на время полу- жизни белка
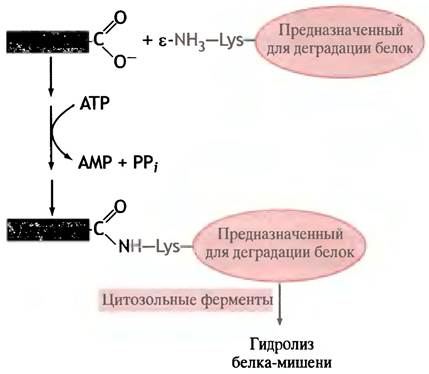
В деградации долгоживущих структурных белков определенную роль играют лизосомы (см. рис. 16.2): они ответственны за аутолитическую деструкцию клеток в ходе развития (например, за исчезновение хвоста у головастиков).
В целом о деградации белка нам известно намного меньше, чем о гораздо более сложном процессе его биосинтеза. А эта область жизненно важна. Вспомним (см. с. 149), что при голодании распад мышечных белков обеспечивает глюконеогенез аминокислотами: без этого человек не смог бы прожить дольше 24 часов из-за полного истощения запасов гликогена. Деградация белка в клетке сейчас активно изучается.
Вопросы к главе 22
1. Почему для кодирования 20 аминокислот используется 61 кодон, а не 64?
2. Типов тРНК меньше, чем 61. Почему?
3. Почему на схемах в спаренном с кодоном состоянии молекулу тРНК располагают в перевернутом виде по сравнению с той же РНК, показанной отдельно?
4. Какие механизмы обеспечивают точность трансляции?
5. Опишите участие и (там, где это известно) роль GТР в синтезе белка.
6. Исследования показали, что у Е. соli молекулы тРНК (с прикрепленными аминоацильными или пептидильными фрагментами) занимают А, Р и Е-участки на рибосоме. Объясните причину и механизм этого явления.
7. Механизм инициации трансляции у эукариот не совместим с полицистронной мРНК. Объясните почему.
8. Объясните роль шаперонов в синтезе белка.
9. Какое заболевание может быть вызвано неправильным фолдингом белка?
10. Объясните, каким образом белки транспортируются через эндоплазматический ретикулум.