БИОХИМИЯ И МОЛЕКУЛЯРНАЯ БИОЛОГИЯ - В. ЭЛЛИОТ - 2002
ГЛАВА 29. РОЛЬ ЦИТОСКЕЛЕТА В ПОДДЕРЖАНИИ ФОРМЫ КЛЕТОК И ОСУЩЕСТВЛЕНИИ МЕХАНИЧЕСКОЙ РАБОТЫ
На первый взгляд кажется, что немышечные клетки механической работы не совершают. Также легко представить животную клетку в виде бесформенного мешка, наполненного протоплазмой. Но и это неправильно, поскольку у клеток есть цитоскелет (комплекс внутренних перегородок), который принимает участие в перемещении внутриклеточных структур, движении клеток и поддержании их формы.
В первом приближении необходимость этих видов механической работы не столь очевидна. Но в таком случае животная клетка без плотной клеточной стенки была бы маленьким аморфным образованием. Форма большинства типов животных клеток индивидуальна, и в ее поддержании важную роль играют внутриклеточные структуры. Например, форма уплощенного двояковогнутого диска эритроцита поддерживается при помощи специальных белковых перегородок, взаимодействующих с мембраной.
Многие клетки постоянно изменяют свою форму, поэтому цитоскелет, определяющий ее, все время изменяется. Такая структура цитоскелета, отличающаяся от структуры, свойственной эритроцитам или микроворсинкам кишечника (см. рис. 4.1), характерна для животных клеток. Растительные и бактериальные клетки с прочными клеточными стенками относятся к другой категории.
Подвижность - неотъемлемое свойство животных клеток; помимо наиболее ярких примеров амебоидного движения макрофагов и других лейкоцитов, мигрирующих в ткани, многие клетки передвигаются по поверхности ткани, что важно в ходе эмбрионального развития и при нормальном заживлении ран. Другой тип активного (немышечного) сокращения наблюдается в процессе деления клетки. Тогда вокруг её экваториальной зоны образуется перетяжка, которая приводит к разделению клетки. Еще одна форма движения - пульсация микроворсинок, способствующая перемещению поверхностных слоев слизи, например, в дыхательных путях, или биение жгутиков сперматозоидов.
Эукариотическая клетка - очень большой объект, если соотносить ее размеры с низкой скоростью диффузии веществ в растворе. Бактериальная клетка намного меньше и поэтому «полагается» на диффузию растворенных веществ, в то время как многие эукариотические клетки нуждаются в сложной внутренней транспортной
системе. Пузырьки аппарата Гольджи должны перемещаться к поверхности клетки или в другие места назначения. Движение молекул мРНК от ядра в цитоплазму также происходит по строго заданному маршруту. Другим ярким примером служит аксон нерва, который может достигать более полуметра в длину; синтез белков и формирование везикул происходит в теле клетки, и некоторые из этих компонентов должны быть доставлены к окончанию аксона. При помощи диффузии такое сделать невозможно! Гигантская клетка водоросли Nitella настолько велика, что диффузия не смогла бы обеспечить ее существование; поэтому клетка поддерживает постоянные транспортные потоки веществ в цитоплазме. Еще одним примером внутриклеточного транспорта может служить расхождение хромосом в дочерние клетки в процессе клеточного деления.
Суммируя все вышесказанное, можно констатировать, что клетки имеют сложную внутреннюю организацию, обеспечивающую поддержание формы, процессы движения и внутриклеточного транспорта. Эта внутренняя организация менее выражена, чем сократительный аппарат мышечных клеток, поскольку: 1) структуры, осуществляющие перечисленные виды работ, выявляются только при определенном окрашивании клеток; 2) многие из них постоянно модифицируются в зависимости от сиюминутных потребностей клетки.
Прежде чем мы перейдем к детальному рассмотрению роли цитоскелета в определении формы клеток и в производстве ею механической работы, стоит отметить ряд важных положений.
✵ Актиновые филаменты есть практически во всех клетках; они участвуют в определении формы животных клеток и в осуществлении амебоидных и «ползающих» движений.
✵ Актиновые филаменты играют также роль внутриклеточных «железнодорожных путей», по которым интенсивно перемещаются составные части клетки.
✵ Миозиноподобные молекулы существуют и в немышечных клетках. Один тип молекул миозина (аналогичный мышечному миозину) осуществляет силовое воздействие на расположенные рядом актиновые филаменты, обеспечивая движение клетки. Другие типы молекул миозина (минимиозины), расположенные вдоль актиновых транспортных путей, перемещают различные внутриклеточные «грузы».
Источником энергии служит АТР.
✵ Особая транспортная система существует в виде микротрубочек, также играющих роль транспортных путей, по которым перемещаются молекулярные «двигатели», тянущие за собой внутриклеточный «груз». Используя энергию АТР, такой «двигатель» воздействует на соседние микротрубочки, производя скользящие движения и выполняя таким образом механическую работу, как и актомиозиновая система.
✵ Третий тип компонентов цитоскелета представлен промежуточными филаментами (они имеют промежуточные размеры по сравнению с размерами филаментов актина и микротрубочек). Одни из них (ламины) формируют сеть, которая является составной частью ядерной оболочки. Промежуточные филаменты придают прочность клеткам эпидермиса, а кератиновые промежуточные филаменты - клеткам волос.
После этого краткого введения рассмотрим компоненты цитоскелета более подробно.
Роль актина и миозина в немышечных клетках
Актин относится к числу простых белков, наиболее широко представленных в большинстве эукариотических клеток. Актины различных типов клеток сходны между собой: это высококонсервативные белки, образующие филаменты по типу описанных для мышечной клетки (см. главу 28). Миозин также является универсальным компонентом эукариотических клеток, хотя и присутствует в них в гораздо меньших количествах. Преобладание актина над миозином в немышечных клетках связано с тем, что помимо участия в сокращении актин играет также структурную роль.
Структурное значение актина и его роль в движении клеток
Наиболее наглядным примером структурной роли актина может служить строение микроворсинок клеток щеточной каемки, пальцевидные выступы которой значительно увеличивают адсорбционную поверхность кишечника (рис. 29.1). Актиновые филаменты пронизывают большинство животных клеток. Особенно плотно они располагаются около цитоплазматической мембраны, в которой закреплены пучки актиновых филаментов, участвующие в поддержании формы клетки (рис. 29.2, а). Упорядоченность актиновых филаментов и образование сетей зависят от множества других белков, связанных с актином. Актиновые филаменты микроворсинок - структуры постоянные, однако и они способны к разборке и самосборке. Когда фагоцит движется в сторону поглощаемой частицы, можно видеть, как актиновые филаменты продолжаются в псевдоподии («лопастеобразные» выросты клетки, вытягивающиеся в направлении движения).
Рис. 29.1. Строение актиновых филаментов микроворсинки. Формирование прочной структуры цитоскелета обусловлено взаимодействием актиновых филаментов с другими белками с образованием поперечных сшивок
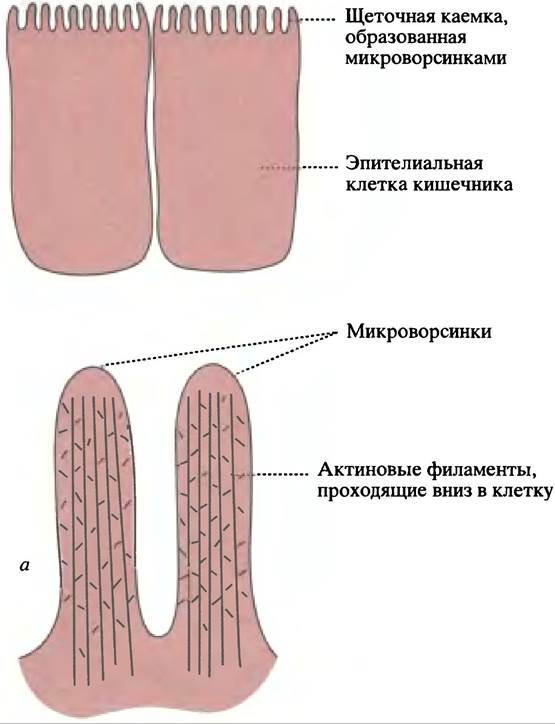
Рис. 29.2. Микрофотографии актиновых волокон и микротрубочек. а - Актиновые волокна миобластов мыши, визуализированные при помощи антител к актину. Волокна пронизывают клетку насквозь и их пучки (волокна напряжения) присоединены в фокальных точках к мембране клетки. Волокна быстро разбираются и собираются. Фотография любезно предоставлена Р. Gunning, Children’s Medical Research Institute, Sydney; б — микротрубочки в цитоплазме клетки, исходящие из центра организации микротрубочек или центросомы
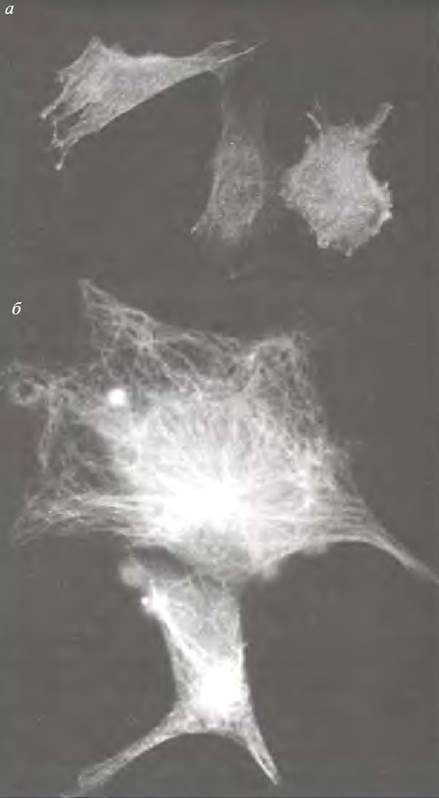
Актиновые волокна участвуют в амебоидных и «ползающих» движениях клеток. Это результат действия миозина II (обычных молекул миозина мышечного типа). Приблизительно 16 таких молекул при необходимости собираются в небольшие биполярные волокна, подобные толстым филаментам поперечнополосатой мышцы. Их действие очень похоже на действие саркомера: биполярные волокна развивают сократительное усилие на соседние актиновые филаменты и вызывают натяжение мембраны клетки, к которой прикреплены филаменты (рис. 29.3). Регуляция сокращения (как и в гладкой мышце) связана с фосфорилированием легких цепей миозина (см. главу 28).
Рис. 29.3. Тянущее действие миозина II на актиновые филаменты. Энергию процесса обеспечивает гидролиз АТР. Выполнение системой полезной работы будет зависеть от соответствующего прикрепления актинового филамента к клеточной мембране для изменения формы клетки. Каждая молекула миозина имеет участок, способный к взаимодействию с другими молекулами, который определяет самосборку биполярного волокна. На схеме представлено продольное сечение - биполярные комплексы имеют цилиндрическую форму. Сокращение обусловлено скольжением актиновых филаментов относительно пучков миозина
Довольно точно определена сократительная роль актомиозиновой системы в цитокинезе - разделении цитоплазмы в процессе деления клетки. Во время анафазы
кольцо актиновых филаментов собирается в экваториальной плоскости, где и происходит разделение. Считается, что актиновые филаменты заякорены в плазматической мембране и перекрывание этих волокон (с противоположной полярностью) дает возможность сокращения цитоплазматическим миозинам. По мере развития сокращения, это образование распадается, и цитоплазма распределяется между двумя дочерними клетками. Такая система служит превосходным примером временной организации сократительных комплексов, входящих в структуру немышечных клеток.
Роль актина и миозина во внутриклеточном транспорте веществ
Актиновые филаменты могут служить транспортными путями, по которым ATP-зависимые молекулярные двигатели перемещают клеточные компоненты. В главе 28 упоминалось, что если в эксперименте отдельные головки миозина (мышечного типа) прикрепить к шарикам латекса, то в присутствии АТР и Са2+ они будут двигаться по иммобилизованным актиновым волокнам. «Хвост» молекулы миозина мышечного типа устроен таким образом, что обеспечивает самосборку миозина в биполярный (толстый) филамент, который способствует скольжению соседних актиновых волокон, вызывая таким образом сокращение.
Существует семейство молекул миозина с обычными миозиновыми головками, но вместо длинного палочкообразного хвоста, свойственного мышечному миозину, у них хвост маленький, отсюда и произошло их название - минимиозины. Молекулы минимиозина не могут формировать биполярные пучки, но за счет энергии АТР они перемещаются по актиновому фила- менту. Хвосты минимиозинов предназначены для присоединения к другим структурам, например, к мембранам везикул: если такое присоединение происходит, минимиозин будет тянуть везикулу по актиновому филаменту. В гигантской водоросли Nitella минимиозины тянут эндоплазматический ретикулум по неподвижным актиновым филаментам через всю клетку. Хвосты минимиозина присоединены к эндоплазматическому ретикулуму, пронизывающему цитоплазму. Они подтягивают его, вызывая непрерывную циркуляцию всего содержимого цитоплазмы.
Идентифицировано множество необычных миозинов (немышечного типа). Они обнаружены как у дрожжей, так и у позвоночных. У всех миозинов немышечного типа есть глобулярные актин-связывающие головки и различные хвосты, которые взаимодействуют с определенными мембранными структурами.
Микротрубочки, клеточное движение и внутриклеточный транспорт
Что такое микротрубочки?
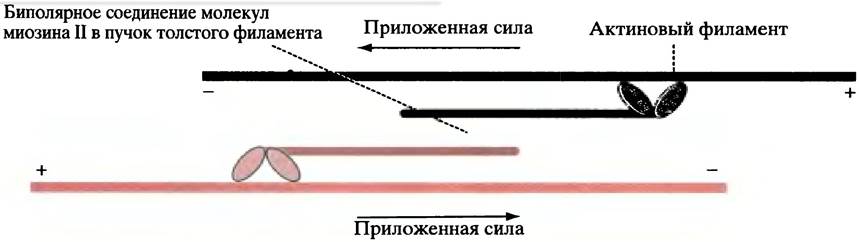
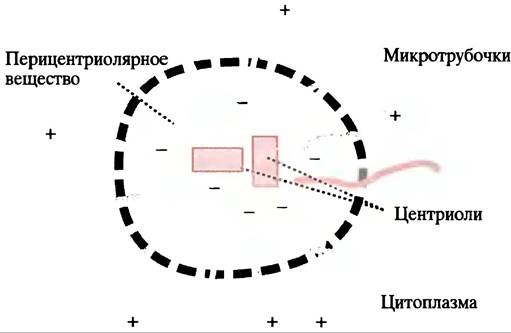
Микротрубочки образуются при полимеризации белка тубулина, который является гетеродимером, образованным субъединицами α- и β-тубулина. В процессе полимеризации α-тубулин одного димера контактирует с β-тубулином следующего с образованием протофиламентов, 13 продольных рядов которых и формируют полую трубку. Полимеризация происходит в направлении от головы к хвосту таким образом, что микротрубочка имеет определенную полярность; ее концы обозначаются соответственно, как (+)- и (-)-концы. Микротрубочки в клетке нестабильны, они могут быстро собираться и разбираться. Нестабильность обусловлена незащищенными концами, которые способствуют быстрому разрушению микротрубочек путем деполимеризации на свободные субъединицы тубулина. В клетке (-)-концы связаны с центром организации микротрубочек (ЦОМТ) - структурой, расположенной около ядра, которая содержит (в животных клетках) пару маленьких телец - центриолей - образованных из слившихся микротрубочек. Клетка пронизана микротрубочками, исходящими из ЦОМТ (см. рис. 29.2, б и 29.4).
Рис. 29.4. Микротрубочки, исходящие из ЦОМТ. Знаки (+) и (-) указывают полярность микротрубочек. Центриоли - пара трубчатых структур, построенных из соединенных микротрубочек; расхождение микротрубочек происходит в веществе, окружающем центриоли
Что защищает (+)-концы микротрубочек?
Как уже отмечалось, микротрубочки быстро распадаются, если их концы не защищены. ЦОМТ защищает (-)-конец. В клетке микротрубочки растут (и распадаются) с (+)-концов. При достижении микротрубочкой соответствующей мишени белки последней защищают ее. Микротрубочки могут расти из ЦОМТ в совершенно случайных направлениях, но лишь те из них, что вступят в контакт с соответствующим компонентом клетки, будут защищены и стабилизированы белками-мишенями, в то время как остальные разрушатся. До сих пор остается неясным, что же защищает от распада растущую микротрубочку, прежде чем она достигнет белковой мишени. Согласно современной модели, субъединица тубулина связывает GTP и в виде такого комплекса защищает (+)-конец. После ее присоединения к трубочке происходит медленный гидролиз GTP до GDP и Рi. Таким образом, вновь добавленные субъединицы тубулина в течение короткого времени будут находиться в комплексе с GTP, что временно защищает конец микротрубочки от разрушения. Гидролиз GTP играет роль уже знакомого нам «молекулярного хрономегражного прибора»: если добавление новых молекул тубулина происходит до того, как последняя присоединенная субъединица потеряет GTP, микротрубочка будет защищена. Если же GTP на свободном конце гидролизуется раньше, то, согласно этой модели, распад микротрубочки неизбежен. В ресничках и жгутиках, где микротрубочки являются постоянной структурой (см. ниже), после их сборки происходит ковалентная модификация белка, препятствующая распаду. Такова основная схема строения микротрубочек. Теперь рассмотрим их функции.
Функции микротрубочек
Как и актиновые филаменты, микротрубочки участвуют в поддержании формы клетки, ее движении и внутриклеточном транспорте.
Молекулярные двигатели, обеспечивающие движение по микротрубочкам
Идентифицированы два типа ATP-зависимых двигателей - кинезин и динеин. Кинезин перемещается по микротрубочке от (-)- к (+)-концу, а динеин - в противоположном направлении. Каждый из них имеет две глобулярные головки, напоминающие миозиновые (рис. 29.5, а), с легкими цепями, направленными в противоположные стороны.
Рис. 29.5. Строение и функция кинезина: а - Строение молекулы кинезина; б - транспортировка везикулы по микротрубочке молекулой кинезина (схематично)
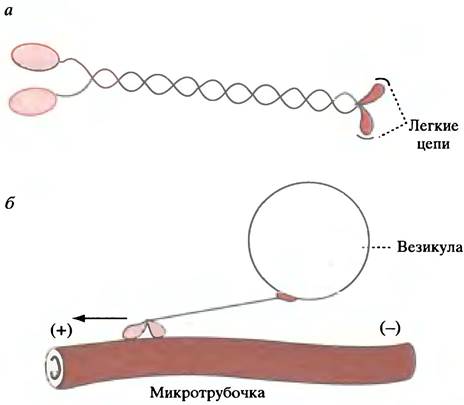
Существуют семейства молекул кинезина и динеина, специализирующиеся на выполнении различных функций в разных тканях. Головки, совершающие движение по микротрубочке, вероятно, не изменяются, а вот структура хвоста модифицируется для соединения с различными структурами клетки (рис. 29.5, б). В качестве примера работы молекулярных двигателей можно рассмотреть перемещение везикулы по микротрубочке (см. ниже). В присутствии АТР молекулы кинезина и динеина (обладающие АТРазной активностью) будут перемещаться по иммобилизованным на твердой поверхности волокнам микротрубочек точно также, как головки миозина передвигаются по иммобилизованным актиновым филаментам.
Роль микротрубочек в движении клеток
Микротрубочки играют важную роль в движении ресничек и жгутиков. Клетки, выстилающие дыхательные ходы, имеют большое количество ресничек, пульсирующее движение которых выводит слизь и удаляет чужеродные частицы. Сперматозоид перемещается при помощи вращения жгутика. Реснички и жгутики
по структуре похожи: они берут начало из базальных телец, напоминающих центриоль, и микротрубочки проходят вниз по всей их длине; к микротрубочкам присоединены молекулы динеина, которые служат двигателями. Используя энергию АТР, они перемещаются по микротрубочкам по направлению к (-)-концу, вызывая смешение последних относительно друг друга и приводя к волнообразным движениям ресничек или жгутиков.
Роль микротрубочек в транспорте везикул внутри клетки
Микротрубочки участвуют во внутриклеточном транспорте. Пузырьки, образуемые аппаратом Гольджи, направляются в различные места клетки строго по назначению. Детали такого запрограммированного движения и перемещения пузырьков неизвестны, но, по-видимому, в нем участвуют микротрубочки и АТР-зависимые двигатели.
Как уже упоминалось, аксон нервной клетки бывает настолько длинным, что его окончание может находиться на значительном расстоянии от тела клетки, где происходит образование везикул и синтез белков. Для их транспорта по аксону вытягиваются микротрубочки, ориентированные (+)-концом к окончанию. Молекулярный двигатель кинезинодним концом присоединяется к мембране пузырька и передвигается по микротрубочке в направлении к (+)-концу (см. рис. 29.5, б).
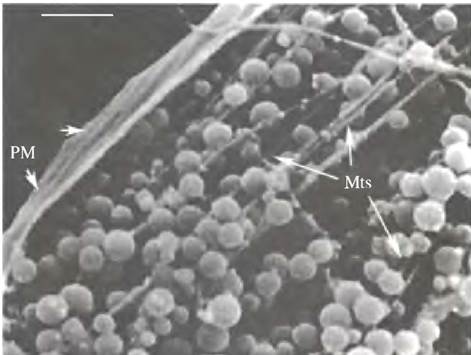
Другой двигатель - цитоплазматический динеин - осуществляет транспорт в обратном направлении от аксона к телу клетки. Наглядным примером движения по микротрубочкам может служить транспорт пигментных везикул (рис. 29.6).
Рис. 29.6 Транспорт пузырьков по микротрубочкам. Микрофотография получена при электронном сканировании транспортируемых по микротрубочкам пузырьков, содержащих пигмент в хроматофоре «рыбы-белки». Изменение цвета клетки вызвано перемещением пузырьков к центру клетки и обратно. Mts - микротрубочки; РМ - плазматическая мембрана. (Размер шкалы = 0,5мкм)
Роль микротрубочек в митозе
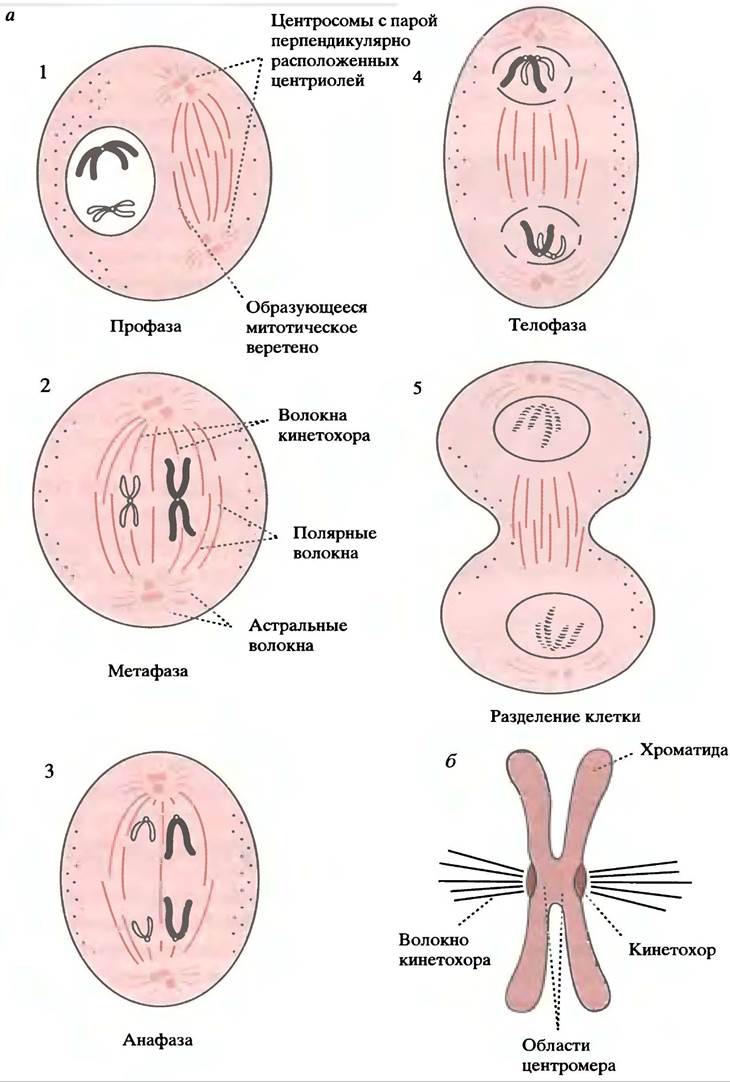
Стадии митотического цикла клетки показаны на рис. 29.7, а.
В профазе центросомы (то же, что и ЦОМТ в неделящихся клетках) удваиваются и расходятся к противоположным полюсам клетки; из них выходят микротрубочки, образуя веретено деления. ДНК уплотняется, формируя набор видимых в световой микроскоп хромосом. В прометафазе ядерная мембрана исчезает, микротрубочки веретена входят в ядерную область и присоединяются к центромерам хроматид. Они представляют собой «сестринские» хромосомы, сформированные при репликации ДНК в интерфазе, которые связаны в центральных областях, образуя Х-образные структуры (см. рис. 19.9). После разделения каждая хроматида станет хромосомой. Хроматиды в метафазерасполагаются в экваториальной плоскости, готовясь к анафазе. Затем хроматиды разделяются и двигаются к полюсам (анафаза А), а те удаляются друг от друга, расходясь в разные стороны (анафаза В). В телофазе происходит восстановление ядерной мембраны. Это сопровождается
разделением клетки путем расхождения цитоплазмы (цитокинез), которое осуществляется актомиозиновой системой. Затем в каждой дочерней клетке центросома становится центром образования множества микротрубочек, характерных для интерфазной клетки.
Механизм перемещения хромосом (анафаза А)
До сих пор неизвестно, что вызывает перемещение хромосом к полюсам клетки после их присоединения к волокнам кинетохора (рис. 29.7, б), однако ясно, что эту работу может совершать молекулярный двигатель типа динеина. В процессе движения хромосом волокна кинетохора укорачиваются. Микротрубочки не способны к сокращению, и поэтому предполагается, что хромосома перемещается по волокну, а микротрубочки подвергаются деполимеризации после ее прохождения.
Механизм разделения полюсов (анафаза В)
В этом процессе участвуют полярные волокна (см. рис. 29.7, а), которые не присоединяются к хромосомам, а перекрываются в экваториальной области. Поскольку полюса перемещаются обособленно, волокна удлиняются, чтобы сохранить это перекрывание. Его наличие позволяет предположить, что в расхождении полюсов участвуют молекулярные двигатели, с которыми связаны астральные волокна, выходящие из полюсов. Независимо от типа используемого механизма, установлено, что разделение полюсов нуждается в АТР.
Рис. 29.7 Митоз: а - Упрощенное изображение стадий митоза. Полярные волокна выделены цветом; предполагается, что они отвечают за расхождение полюсов в разные стороны. Черные волокна - волокна кинетохора, ответственные за перемещение хромосом к полюсам. Все волокна являются микротрубочками; б - присоединение волокна кинетохора (схематично)
Промежуточные филаменты
Описанная выше актиновая сеть содержит микрофила- менты диаметром 6 нм; диаметр микротрубочек 20 нм. Третий тип сети в клетках эукариот образован филаментами с диаметром в среднем 10 нм, поэтому их называют промежуточными филаментами (ПФ).
Существует несколько типов ПФ, образованных различными группами белков, которые характеризуются высокой гомологичностью. Обычно они имеют стержневой филамент, содержащий приблизительно 350 аминокислотных остатков с изменяющимися у разных типов ПФ-концами. Это разнообразие белков контрастирует с однотипными белковыми субъединицами актиновых филаментов и микротрубочек.
Различные типы ПФ, обнаруженные в клетках эукариот, экспрессируются на различных стадиях развития и дифференцировки. Впрочем, роль сетей ПФ остается невыясненной. Как уже упоминалось, промежуточные филаменты придают прочность клеткам эпидермиса.
Белковые ламины образуют сеть, соединенную с внутренней поверхностью внутренней ядерной мембраны; они связаны с ядерными порами. В нервных клетках существуют нейрофиламенты - механическая опора длинным аксонам; филаменты десмина расположены в Z-дисках саркомеров. Вызывает недоумение тот факт, что недостаток промежуточных филаментов в культивируемых клетках, вызванный мутациями или введением определенных антител, не мешает им расти и благополучно делиться.
Вопросы к главе 29
1. Какие функции выполняет актин в немышечных клетках?
2. Что такое микротрубочки?
3. Микротрубочки с незащищенными концами подвергаются спонтанной диссоциации. Что защищает их концы?
4. Что такое кинезин и динеин? Проиллюстрируйте ответ схемой.
5. В процессе деления клетки в метафазе хромосомы движутся от экваториальной плоскости обособленно. Микротрубочки присоединены к кинетохорам и сокращаются по мере расхождения хромосом. Означает ли это, что микротрубочки сокращаются? Объясните ответ.
6. Что такое промежуточные филаменты?
7. Каковы функции промежуточных филаментов?