Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Молекулярная генетика и системы регуляции
Молекулярная генетика
Процессы экспрессии генов
Мы уже знаем, что молекула ДНК представляет собой очень длинную двойную спираль, состоящую из двух цепей. Обе цепи представляют собой полимеры, построенные из четырех нуклеотидов, отличающихся друг от друга природой азотистого основания [аденина (А), гуанина (G), цитозина (С) и тимина (Т)]. Две цепи в двойной спирали ДНК имеют комплементарные последовательности так, что А в одной цепи всегда связывается с Т другой; аналогично связываются G и С.
Геном называют участок ДНК, кодирующий определенный белок. Экспрессия гена, в результате которой формируется аминокислотная последовательность соответствующего белка, представляет собой двустадийный процесс; промежуточным продуктом этого процесса является матричная (или информационная) рибонуклеиновая кислота (мРНК или иРНК). На первом этапе экспрессии гена, называемом транскрипцией, сложный олигомерный фермент РНК-полимераза катализирует синтез мРНК на матрице, роль которой выполняет ген. На втором этапе экспрессии гена на основе содержащейся в нуклеотидной последовательности мРНК информации синтезируется пептидная цепь с соответствующей аминокислотной последовательностью. В этом процессе, называемом трансляцией, участвуют различные компоненты клетки, в том числе центры и регуляторы пептидного синтеза — рибосомы, а также несколько различных транспортных РНК (тРНК), которые переносят химически активированные аминокислоты к месту синтеза пептидной цепи в соответствии с диктуемой мРНК последовательностью. Ниже мы рассмотрим эти процессы, а также генетический код, являющийся связующим звеном между нуклеотидной и аминокислотной последовательностями. В нашем обсуждении мы будем рассматривать только основные моменты и по необходимости опустим немаловажные в ряде ситуаций детали; поэтому читателю настоятельно рекомендуется подробнее ознакомиться с проблемой экспрессии генов в приведенной в конце главы литературе.
Транскрипция начинается со связывания фермента РНК-полимеразы с сигнальной нуклеотидной последовательностью, так называемым промотором, располагающейся на одной из комплементарных цепей ДНК. Связанная с РНК-полимеразой нуклеотидная цепь становится таким образом матрицей ДНК для синтеза молекулы мРНК. После частичного расплетания двойной спирали ДНК вблизи от комплекса РНК-полимеразы с промотором полимераза начинает двигаться вдоль матричной цепи ДНК в направлении 3'→5'. При построении комплементарной ДНК последовательности одноцепочечной молекулы мРНК соблюдается правило образования пар нуклеотидов ДНК—РНК (вспомните разд. 2.3.2 или см. рис. 6.1). Синтезируемая мРНК располагается антипараллельно матричной цепи ДНК; следовательно, мРНК синтезируется от 5'-конца к 3'-концу. В приведенном на рис. 6.1 примере матрицей является нижняя цепь ДНК. Транскрипция прекращается, т. е. синтез мРНК останавливается в тот момент, когда РНК-полимераза достигает специфической нуклеотидной последовательности матричной цепи (терминатора), сигнализирующей об окончании синтеза. Прежде чем перейти к рассмотрению следующих стадий, следует отметить, что РНК-полимераза связывается с ДНК и неспецифично в центрах, не являющихся промоторами. Неспецифично связанная полимераза не инициирует и не осуществляет транскрипцию.

РИС. 6.1. Схематическое изображение транскрипции матричной цепи ДНК (здесь транскрибируется нижняя цепь) при участии фермента РНК-полимеразы, в результате чего синтезируется молекула соответствующей мРНК с комплементарной нуклеотидной последовательностью.
Длина молекулы мРНК изменяется от приблизительно 300 до 3000 нуклеотидных остатков и более. Обычно одна мРНК соответствует одному гену в цепи ДНК, но в некоторых случаях молекула мРНК несет генетическую информацию о группе родственных и тесно связанных генов — так называемом опероне (см. разд. 6.1.4). Как мы уже отмечали при обсуждении энергетики биосинтеза в гл. 5, построение цепи РНК осуществляется путем отщепления пирофосфатной группировки от нуклеозидтрифосфатов. Следовательно, образование каждой фосфодиэфирной связи РНК сопровождается расщеплением двух высокоэнергетических фосфатных связей. Это способствует тому, что реакция конденсации протекает «до конца» и свидетельствует о важности синтеза этих биополимеров для нормального функционирования клетки.
Прежде чем перейти к описанию процесса трансляции, нам нужно несколько более детально ознакомиться с транспортными РНК. Известно, что каждая разновидность тРНК может транспортировать только одну соответствующую ей аминокислоту. На рис. 6.2 изображена вторичная структура аланил-тРНК из дрожжей. Присоединение аланильного остатка к соответствующей тРНК требует предварительной активации аланина путем его превращения в аланил-АМР, как это отражено в уравнении реакции (5.39). Активированный аланиновый остаток затем связывается с соответствующей тРНК при участии фермента, специфичного как в отношении аминокислоты, так и в отношении тРНК. Таким образом обеспечивается связывание аланина именно с аланиновой тРНК.
Теперь мы хможем перейти к обсуждению природы специфичности тРНК. В основании нижней петли молекулы тРНК имеется последовательность из трех нуклеотидов, называемая антикодоном. Этот элемент структуры присутствует во всех изученных тРНК. Последовательность оснований антикодона комплементарна трехнуклеотидному сегменту мРНК, называемому кодоном. Изучение биохимии нуклеиновых кислот показало, что каждый кодон представляет собой «слово» в генетическом послании, т. е. каждый кодон соответствует одной аминокислоте. Поскольку химический алфавит РНК состоит всего лишь из четырех букв (четырех оснований А, С, G и U), а всего должно быть не менее 20 «слов» (по числу аминокислот), то для передачи генетической информации должен существовать специальный «язык» или код. Как это ни удивительно, но генетический код был полностью расшифрован; более того, оказалось, что он практически универсален для всех живых организмов — бактерий, растений и животных (рис. 6.3).
Процесс трансляции начинается со связывания меньшей из двух субъединиц рибосомы с молекулой мРНК с участием специфической последовательности, называемой связывающим рибосому сайтом. Этот сайт обычно расположен на определенном расстоянии, различном у прокариот и эукариот, «выше» кодона AUG, с которого почти всегда начинается считывание информации о последовательности полипептидной цепи. Здесь термин «выше» означает, что этот сайт расположен в направлении 5'-конца от кодона AUG. Последовательность кодонов, определяющая аминокислотную последовательность белковой цепи, начинается с кодона AUG и идет в направлении 5'→3'.

РИС. 6.2. Антикодон аланиновой тРНК узнает комплементарный кодон из трех оснований в мРНК. (Уотсон Дж., Молекулярная биология гена. — М.: Мир, 1978.)
Далее мРНК образует комплекс со всей рибосомой, и к этому комплексу присоединяются тРНК, несущие аминокислоты, которые отвечают двум первым кодонам мРНК. В соответствии) с заданной аминокислотной последовательностью между карбоксильной группой первой аминокислоты и аминогруппой второй образуется пептидная связь. Затем рибосома смещается в направлении 3' на один кодон; при этом высвобождается тРНК первой аминокислоты, после чего с комплексом связывается третья аминокислота в виде соответствующей аминоацил-тРНК и присоединяется к пептидной цепи, в результате чего последняя удлиняется еще на один остаток. Этот процесс удлинения полипептидной цепи (рис. 6.4) продолжается вплоть до терминирующего кодона (см. рис. 6.3); в этот момент пептидная цепь отделяется, а комплекс рибосомы и мРНК разрушается. Таким образом, построение полипептидной цепи начинается с N-концевого остатка и завершается С-концевым аминокислотным остатком.
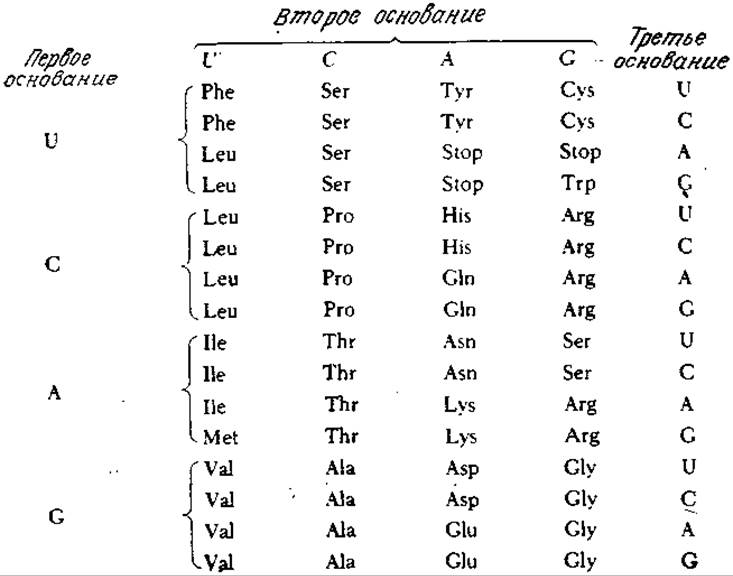
РИС. 6.3. Генетический код. Три кодона (Stop) сигнализируют о прекращении синтеза пептидной цепи. Нетрудно видеть, что замена одного основания кодона (особенно третьего) часто приводит к близкой аминокислоте (принцип надежности).
На рис. 6.5 процесс трансляции изображен с другой точки зрения; здесь основное внимание уделено передаче информации от нуклеотидной последовательности мРНК к соответствующей аминокислотной последовательности полипептида. Образовавшаяся в результате транскрипции (рис. 6.1) гипотетическая мРНК транслируется в соответствующий полипептид. Обратите внимание на то, что здесь тринуклеотидная последовательность AUG выполняет две функции, указывая, во-первых, на начало кодирующей последовательности, для белка и, во-вторых, кодируя метионин (Met) как N-концевой аминокислотный остаток полипептида. Заметьте также, что не вся молекула мРНК несет в себе информацию о структуре синтезируемого белка. Выше мы уже упоминали о предшествующем участку копирования связывающем рибосому сайте; другие нуклеотидные последовательности, в том числе кодирующие последовательности со своими инициирующими и терминирующими кодонами, могут быть расположены ниже (в направлении 5' → 3') кодирующей области.
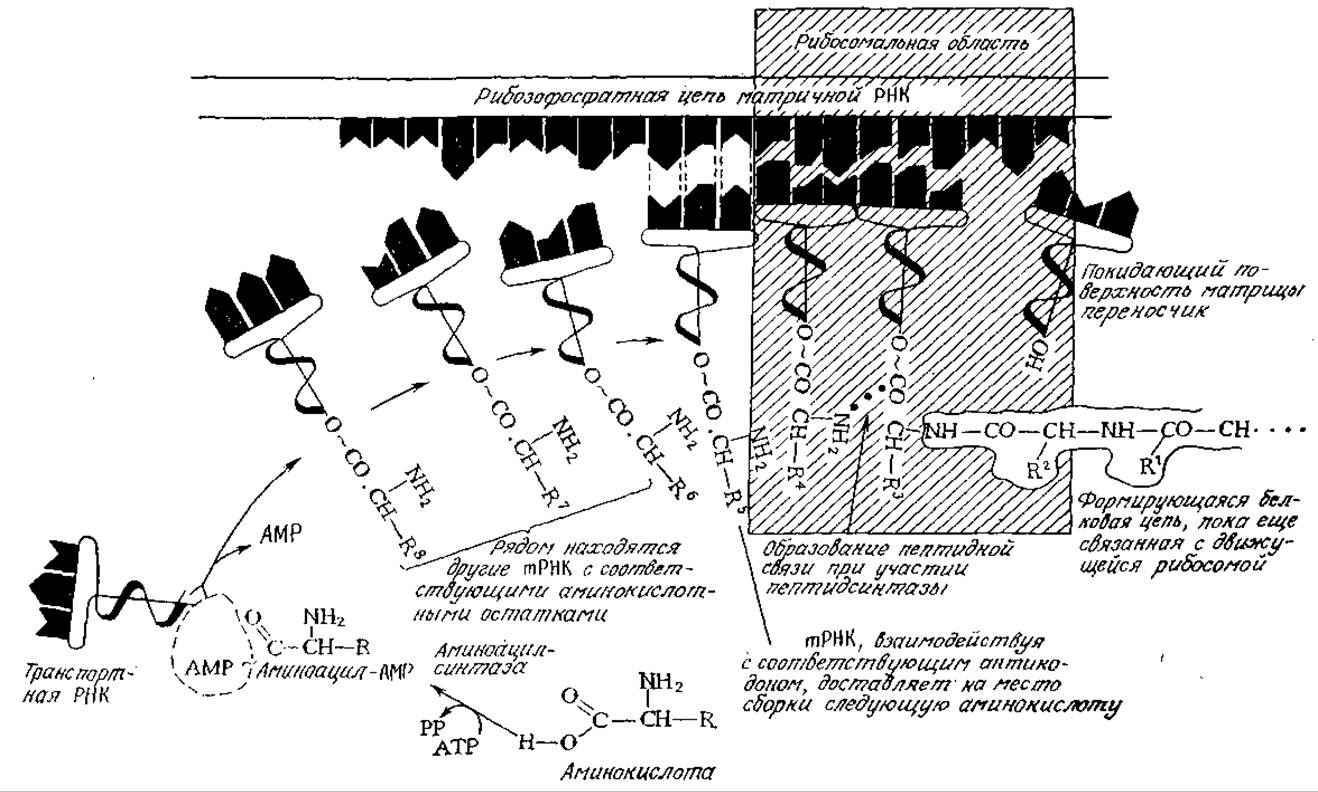
РИС. 6.4. Трансляция — реализация генетической информации мРНК в первичной структуре белка — осуществляется путем ряда специфических взаимодействий между мРНК и тРНК на рибосоме. (Воспроизведено из работы: Edwards N. А., Нassall К. А., Cellular Biochemistry and Physiology, p. 342, McGraw-Hill Publishing Company Ltd., London, 1971.)

РИС. 6.5. Схема передачи (трансляции) информации, содержащейся в нуклеотидной последовательности мРНК, аминокислотной последовательности соответствующего полипептида на рибосоме. Здесь изображена та же мРНК, что и на рис. 6.1.
Здесь полезно рассмотреть процесс экспрессии гена также с точки зрения регулирующих этот процесс элементов, поскольку контролирование последних немаловажно для успешного решения задач генетической инженерии. На рис. 6.6 в самом общем виде приведены важнейшие функции регуляторных последовательностей в процессе передачи генетической информации. Слева и справа от гена расположены последовательности промотора и терминатора, контролирующие начало (инициацию) и окончание (терминацию) транскрипции. ДНК должна содержать также регуляторные сигналы трансляции (связывающий рибосому сайт, кодон-инициатор и терминирующий кодон), которые должны транскрибироваться в мРНК. Наконец, часть мРНК, комплементарная структурному гену ДНК, который кодирует определенную аминокислотную последовательность, должна использоваться для направления белкового синтеза в ходе трансляции.
Часто одна мРНК связывается с несколькими рибосомами. Полирибосомы (полисомы), представляющие собой сложные агрегаты из нескольких индивидуальных рибосом, часто обладают высокоупорядоченной структурой. В полирибосомах одновременно осуществляется считывание генетической информации с различных участков мРНК, что существенно ускоряет белковый синтез и обеспечивает высокую эффективность этого процесса.
Как мРНК, так и тРНК, и рибосомальные рибонуклеиновые кислоты (рРНК) синтезируются на матрице — определенном участке цепи ДНК. Таким образом, ДНК содержит в закодированном виде информацию не только о первичной структуре белков, но и о строении различных веществ, с помощью которых осуществляется белковый синтез.

РИС. 6.6. Регулирующие и информационные последовательности ДНК направляют процессы транскрипции и трансляции (экспрессию гена).
Здесь полезно напомнить, что взаимодействия между аминокислотными остатками полипептидной цепи определяют вторичную, третичную и даже четвертичную структуры белков (разд. 2.4). Следовательно, слабые взаимодействия между атомами и группами играют большую роль на всех этапах хранения генетической информации, ее передачи и реализации.
Прежде чем перейти к вопросу о механизмах передачи генетической информации от поколения к поколению и о факторах, нарушающих этот процесс, следует упомянуть о различиях в механизмах синтеза белков в эукариотах и прокариотах. В клетках бактерий синтез мРНК и трансляция мРНК в белки на рибосомах осуществляются почти в одном и том же центре нуклеоида. В эукариотах транскрипция происходит в ядре. Затем молекулы мРНК претерпевают модификацию (описанную в следующем разделе), диффундируют через поры ядерной мембраны в цитоплазму и образуют комплексы с рибосомами, где и происходит процесс трансляции. В эукариотах и в меньшей степени в прокариотах продукт трансляции часто претерпевает модификацию, результатом которой является окончательная, функционально-активная молекула белка. Посттрансляционные модификации белков рассматриваются в разд. 6.1.3. Наконец, следует отметить, что в эукариотах и прокариотах процесс синтеза белка различно протекает и во времени; подробнее этот вопрос мы рассмотрим в разд. 6.5.