Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Молекулярная генетика и системы регуляции
Молекулярная генетика
Расщепленные гены и модификация мРНК в эукариотах
Матричная РНК эукариот модифицируется путем присоединения к 5'-концу особого структурного элемента, называемого кэпом или кэп-структурой (остаток 7'-метилгуанозина); кэп-структура повышает устойчивость мРНК к действию фосфатаз и нуклеаз. Кроме того, к 3'-концу большинства мРНК эукариот присоединяется последовательность поли-А, содержащая от 150 до 200 остатков дезоксиаденилата. Этот факт можно использовать при выделении эукариотических мРНК, сорбируя их на носителях, содержащих ковалентно-связанные олигонуклеотидные цепи поли-Т.
В настоящее время можно считать твердо установленным, что многие эукариотические гены представляют собой чередование последовательностей, кодирующих структуру белка и не несущих информацию о белковой структуре. Последовательности первого типа иногда называют экзонами, а некодирующие последовательности — интронами или промежуточными последовательностями. Последние, как следует из их названия, располагаются между экзонами и разделяют кодирующие последовательности участками, которые на первый взгляд содержат ненужную избыточную информацию. Очевидно, что в эукариотических клетках должны существовать механизмы, распознающие экзоны и интроны и обеспечивающие в процессе экспрессии гена непосредственную трансляцию одних только экзонов.
Основные структурные элементы эукариотического гена и главные этапы его экспрессии изображены на рис. 6.7. В верхней части рисунка расположен гипотетический сегмент структурного эукариотического гена, содержащий экзоны и интроны. Во время транскрипции матричная цепь ДНК направляет синтез мРНК (первичного транскрипта), комплементарной всей последовательности матрицы, т. е. содержащей последовательности как экзонов, так и интронов.
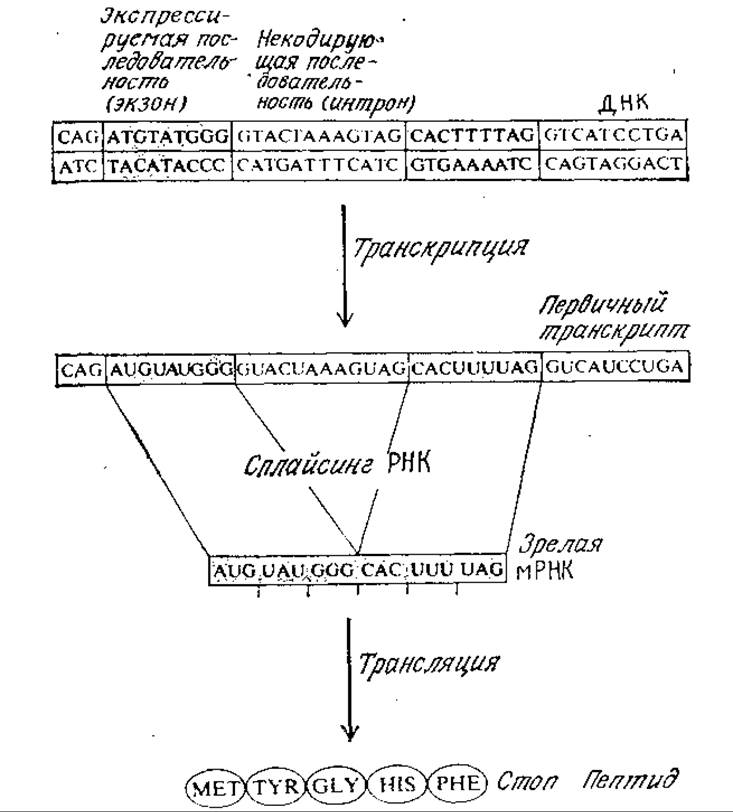
РИС. 6.7. В процессе экспрессии генов эукариот первичный транскрипт мРНК. подвергается сплайсингу; в результате образуется зрелая (активная) мРНК, участвующая в трансляции. Здесь изображены гипотетические укороченные последовательности.
Далее следует характерная, по всей вероятности только для эукариот, стадия сплайсинга. В процессе сплайсинга участки мРНК, отвечающие интронам, вырезаются, а оставшиеся последовательности, комплементарные последовательностям экзонов, соединяются в том же порядке, в каком они располагались в первичном транскрипте. Образующаяся зрелая мРНК затем транслируется описанным выше путем, в результате чего синтезируется соответствующий данному гену белок.
Имеющиеся в настоящее время экспериментальные данные говорят о том, что почти все эукариотические гены содержат интроны. Впрочем, известно и несколько исключений из этого правила, в том числе гены а-интерферона человека. Интроны могут быть очень большими; возможно, что в состав интронов входит более половины всех нуклеотидных остатков ДНК.
Ко времени написания настоящей главы биологическая функция интронов оставалась невыясненной. Возможно, что интроны участвуют в регуляции экспрессии эукариотических генов. С точки зрения эволюции организмов интроны могут выполнять роль инструмента для осуществления относительно крупных изменений генов посредством включения или исключения последовательности целого интрона или экзона. Предполагалось также, что экзоны кодируют довольно крупные участки (домены) первичной структуры белков и что перестановка экзонов дает клетке возможность синтезировать новые белки и таким путем оценивать их функциональную пригодность. С технологической точки зрения с нитронами связаны известные затруднения при экспрессии эукариотические генов в прокариотических клетках-хозяевах, поскольку последние не обладают аппаратом сплайсинга РНК. В разд. 6.3, посвященном технологии рекомбинантных ДНК, мы увидим, как можно преодолеть это затруднение.