Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Молекулярная генетика и системы регуляции
Рост и самовоспроизведение изолированной клетки
Клеточный цикл Е. coli
Простая прокариота Е. coli размножается бинарным делением; в богатой питательными веществами среде число клеток этого организма может удваиваться менее чем за 20 мин. Бактерии Е. coli имеют палочкообразную форму, длина которой может служить надежной мерой объема и массы клетки. Непосредственным изучением изменения длины отдельных клеток Е. coli под микроскопом показано, что в процессе клеточного цикла длина бактерий возрастает постоянно; зависимость длины клетки от времени удовлетворительно описывается возрастающей экспоненциальной функцией. В течение клеточного цикла аналогично изменяется и суммарное содержание всех белков.
В то же время содержание ДНК в течение клеточного цикла Е. coli изменяется довольно сложным образом; по характеру этой зависимости Е. coli резко отличается от эукариотических клеток. Координация процессов синтеза ДНК и деления клеток в случае Е. coli зависит от ряда регуляторных и кинетических параметров, которые мы и рассмотрим ниже.
В самых различных условиях роста, когда деление осуществляется через различные промежутки времени, период репликации хромосомы Е. coli остается приблизительно постоянным (около 40 мин). В процессе синтеза ДНК* скорость движения репликационной вилки (участка молекулы ДНК, в котором осуществляются разделение цепей и синтез ДНК) также остается постоянной. Следовательно, скорость синтеза ДНК прямо пропорциональна числу активных репликационных вилок. Репликация ДНК всегда начинается в определенном центре хромосомы, называемом участком начала репликации. Две вилки движутся в противоположных направлениях от участка начала репликации вокруг циклической хромосомы Е. coli и сходятся в момент окончания процесса репликации.
Регуляция процесса деления клеток связана с синтезом ДНК; бактериальная клетка делится примерно через 20 мин после того, как репликационные вилки достигают конца хромосомы. Как показано на рис. 6.32, ситуация в целом остается сравнительно простой, если Е. coli растет в среде, позволяющей клеткам делиться в среднем раз в час. В этом случае синтез ДНК осуществляется с постоянной скоростью в течение первых сорока минут клеточного цикла, и в оставшиеся двадцать минут, предшествующие делению, клетка не синтезирует ДНК. Росту клетки Е. coli с пятидесятиминутным клеточным циклом (например, при слегка повышенной температуре) отвечает более сложная картина синтеза ДНК; в этом случае новый набор репликационных вилок начинает синтез ДНК за 10 мин до деления клетки. Следовательно, в течение последних 10 мин этого клеточного цикла ДНК синтезируется вдвое быстрее, чем в любой момент медленного (часового) цикла. Нетрудно видеть, что кинетика синтеза ДНК в Е. coli и, вероятно, в некоторых других бактериях в общем случае довольно сложна и характеризуется чередованием периодов спада и повышения синтетической активности; характер чередования зависит от возраста клетки и продолжительности клеточного цикла.
Относительно зависимости синтеза РНК от времени клеточного цикла имеется немного данных. Предполагается, что синтез РНК осуществляется непрерывно. В то же время показано, что для фракции РНК, представляющей собой, по-видимому, мРНК, характерны флуктуации. Следовательно, и скорость синтеза индивидуальных белков может изменяться циклически.
* В настоящем разделе под синтезом ДНК мы подразумеваем только синтез ДНК хромосомы; репликацию плазмид мы рассмотрим позднее.
экспериментальных работ, посвященных главным образом изучению ферментов, активность которых может быть индуцирована или подавлена. Синтез большого числа индуцибельных ферментов в бактерии можно стимулировать в любой момент клеточного цикла. В синхронной культуре скорость синтеза индуцибельных ферментов изменяется ступенчато, что, вероятно, обусловлено удвоением числа соответствующих этим ферментам
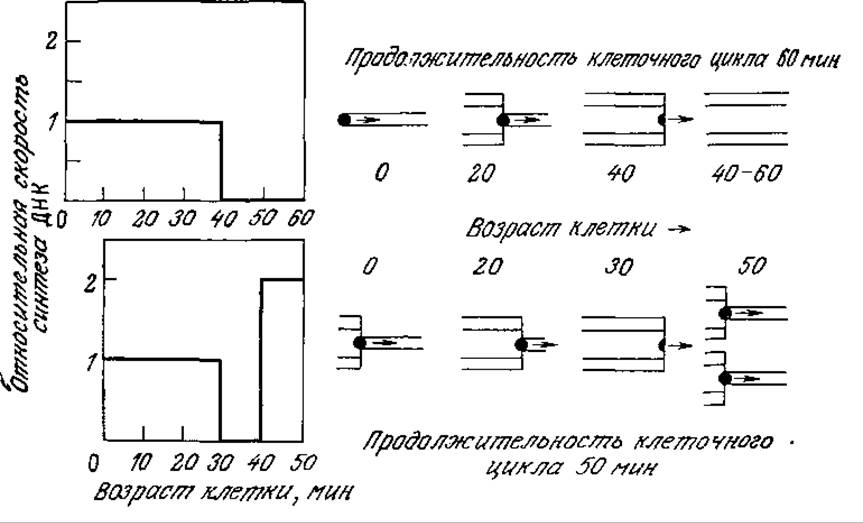
РИС. 6.32. Зависимость скорости синтеза ДНК и структуры хромосомы в Е. coli от возраста клетки и продолжительности периода ее удвоения. В правой части рисунка схематично изображена половина хромосомы Е. coli от участка начала репликации до участка завершения репликации. Черными точками обозначены репликационные вилки.
Такое поведение и в самом деле было обнаружено в ряде генов в момент репликации ДНК. В случае репрессии ферментов имеется несколько возможностей. Если бактерии растут в условиях, когда конечный продукт участка метаболического пути подавляет синтез фермента в максимальной или в минимальной степени, то скорость синтеза фермента остается постоянной в течение промежутка времени, равного продолжительности клеточного цикла, а затем возрастает вдвое. Для промежуточных уровней подавления характерно регулярное периодическое изменение скорости синтеза фермента. Некоторые экспериментальные данные позволяют предположить, что эти периодические изменения обусловлены устойчивыми колебаниями в системе регуляции, функционирующей по принципу обратной связи.
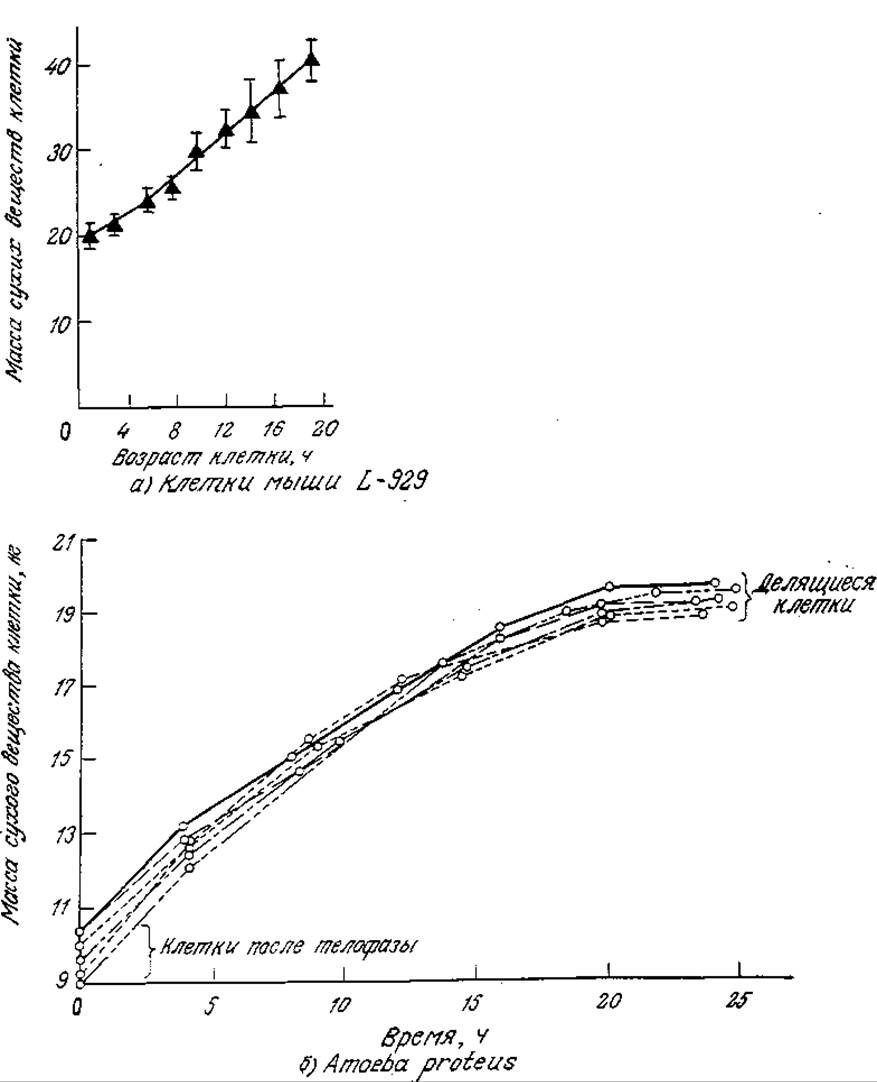
РИС. 6.33. а — Увеличение массы клетки ткани мыши в течение клеточного цикла. [Воспроизведено из статьи: Killander D., Zetterberg А., Quantitative Cytochemical Studies on Interphase Growth. I. Determination of DNA, RNA and Mass Content of Age Determined Mouse Fibroblasts in vitro and of Intercellular Variation in Generation Time, Exp. Cell Res., 38, 272 (1965).] б — рост массы Amoeba proteus в течение клеточного цикла; каждая кривая отвечает одной из шести различных индивидуальных клеток. [Воспроизведено из статьи: Prescott М., Relations Between Cell Growth and Cel Division. I. Reduced Weight, Cell Volume, Protein Content, and Nuclear Volume of Amoeba proteus from Division to Division, Exp. Cell Res., 9, 328 (1955).]