Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Химические основы жизни
Биохимические соединения смешанного строения
Клеточные стенки; пептидогликаны и липополисахариды
В разделе, посвященном липидам, мы уже говорили о том, что клеточные мембраны играют жизненно важную роль в регуляции транспорта веществ в клетку и из клетки. В этом отношении не менее важны и другие структурные элементы наружных поверхностей клеток микроорганизмов и тканей. С точки зрения условий существования микроорганизмы, например бактерии, живут в гораздо более изменчивом и менее контролируемом окружении, чем клетки тканей животных, например клетки печени, поэтому микроорганизмы должны обладать гораздо большей жесткостью, устойчивостью к физическим нагрузкам и резким изменениям осмотического давления. Отсюда следует, что по структуре наружные оболочки бактерий должны сильно отличаться от клеток животных.
В процессах биохимической технологии оболочки клеток представляют интерес в нескольких аспектах. Во-первых, свойства наружных поверхностей клеток определяют их способность к адгезии друг с другом, а также со стенками реакторов, трубопроводов и сепараторов. Как мы увидим в последующих главах, такие явления необходимо учитывать при получении иммобилизованных клеточных катализаторов. Они влияют также на работу микробных реакторов непрерывного действия и на процессы отделения клеток от жидкости. Химические и механические характеристики оболочек определяют также устойчивость клетки к воздействию физических, ферментативных и химических факторов, что существенно в процессах выделения компонентов клетки.

РИС. 2.24. Схематическое изображение структуры стенок грамположительных и грамотрицательных бактерий. Обозначения: ЦМ — цитоплазматическая мембрана; ПГ — пептидогликан; ПП — периплазматическое пространство; ВМ — внешняя мембрана. (Воспроизведено из работы: Sianier R. Y., Adelberg Е. А., Ingraham J. L., Wheelis М. L., Introduction to the Microbial World, p. 128, Prentice-Hall, Englewood Cliffs, N.Y. 1979.)
Как уже упоминалось выше при обсуждении реакции Грама, структуры оболочек грамположительных и грамотрицательных клеток сильно отличаются друг от друга (рис. 2.24). У тех и у других оболочки состоят из нескольких слоев, но их положение, толщина и состав далеко не идентичны. На этом рисунке не показаны различные белки, находящиеся внутри клеточной мембраны или закрепленные на ее поверхности; их мы рассмотрим ниже, в гл. 5, в ходе изучения транспорта веществ через мембраны.
Грамположительные бактерии имеют одну мембрану, а грамотрицательные клетки — две аналогичные мембраны. Между наружной и цитоплазматической мембранами в грамотрицательных бактериях находится область, называемая периплазматическим пространством или периплазмой. В периплазматическом пространстве, включающем от 20 до 40 процентов общей массы клетки, находится ряд ферментов, а также белков, связывающих сахара и аминокислоты.
Как в грамположительных, так и в грамотрицательных бактериях непосредственно к внешней поверхности цитоплазматической мембраны примыкает пептидогликановый слой. Пепти- догликаны построены из остатков дисахарида [состоящего из N-ацетилмурамовой кислоты (NAM) и N-ацетилглюкозамина (NAG), связанных β-1,4-гликозидной связью], мостикового пентапептида, состоящего только из остатков глицина, и тетрапептида. Строение последнего зависит от вида микроорганизма; в Staphylococcus aureus, например, этот тетрапептид содержит остатки L-аланина, D-глутамина, L-лизина и D-аланина (рис. 2.25, а; обратите внимание на наличие редких D-аминокислот). Все перечисленные элементы структуры пептидогликанов связаны множеством поперечных связей, образуя как бы одну гигантскую макромолекулу, окружающую всю клетку (рис. 2.25,6).
Фермент лизоцим, структура которого приведена на рис. 2.17, является эффективным антибактериальным агентом. В основе его бактерицидного действия лежит гидролиз гликозидных ß-1,4-связей между остатками NAM и NAG пептидогликана, приводящий к разрушению и удалению пептидогликановой оболочки бактерий и таким путем к разрыву (лизису) клетки в нативных гипотонических растворах. В лабораторных условиях в изотонической среде удается получить живые клетки, не содержащие пептидогликановых оболочек. После обработки лизоцимом в таких условиях образуются клетки, которые теряют свойственные им очертания и принимают сферическую форму; их называют сферопластами. Если известно, что клеточная стенка удалена полностью, то такие клетки называют протопластами.
Важными компонентами внешней части наружных мембран грамотрицательных бактерий являются липополисахариды. Некоторые липополисахариды называют эндотоксинами, поскольку они в высшей степени токсичны для животных. Токсичность липополисахаридов — одна из причин того, почему заражение крови Е. coli может быть чрезвычайно опасным. По этой же причине при очистке белков, синтезируемых генетически трансформированной Е. coli, необходимо тщательное удаление эндотоксинов.
Молекулы липополисахаридов внешней мембраны имеют три участка: а) липидный компонент А, состоящий из шести ненасыщенных жирных кислот, углеводородные цепи которых проходят в мембрану; они связаны с остатком диглюкозамина; б) центральную олигосахаридную область, построенную из остатков десяти моносахаридов; некоторые из них относятся к числу редких сахаров; в) боковую О-цепь, состоящую из множества повторяющихся тетрасахаридных остатков. Центральная область и О-цепь направлены от клетки в среду. Таким образом, именно наружные боковые О-цепи взаимодействуют с иммунной системой зараженного животного. Путем мутаций бактерии могут достаточно быстро менять структуру О-цепей, что является частью системы их антииммунной защиты.
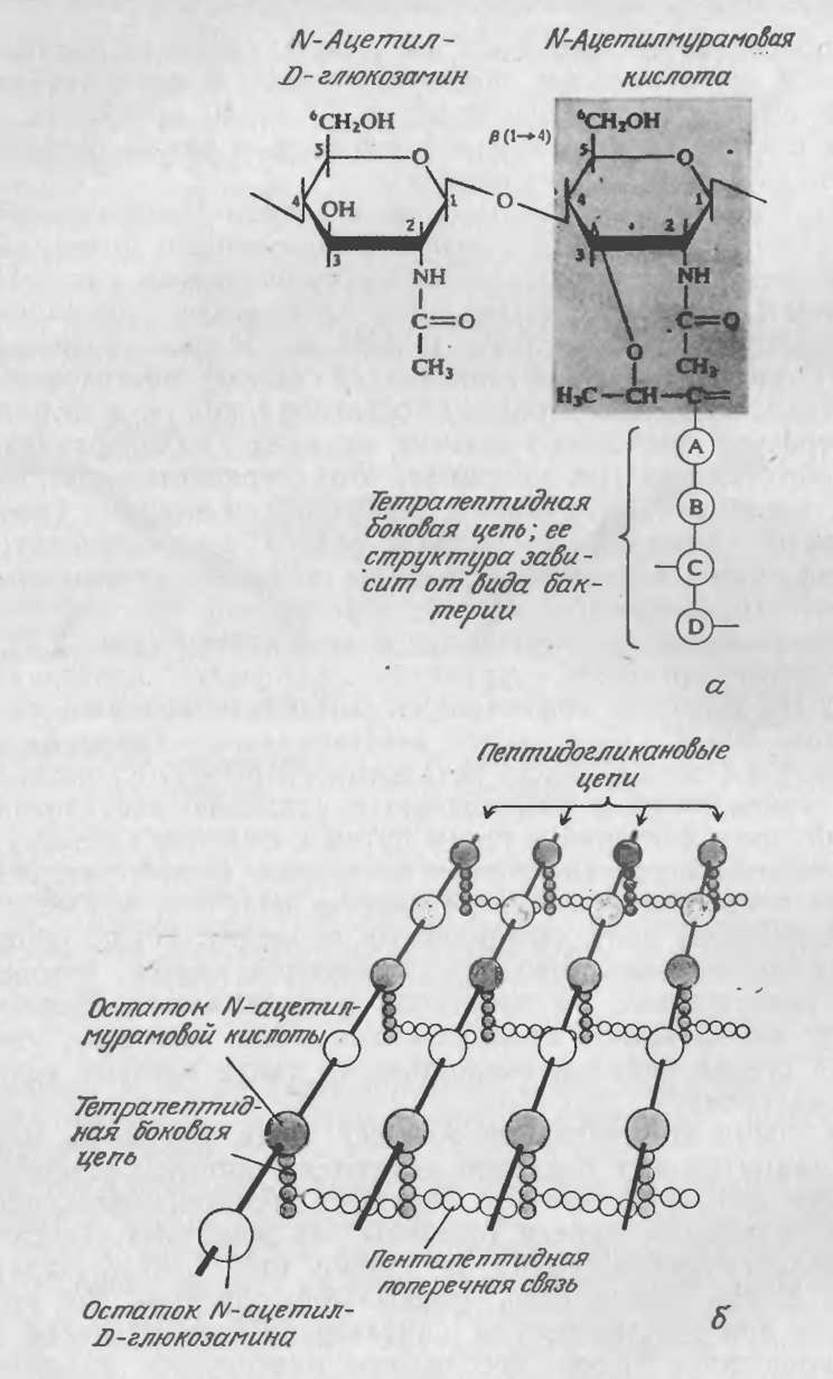
РИС. 2.25. а — строение основных структурных элементов пептидогликанов; б — схематическое изображение пептидогликана клеточной стенки грамположительной бактерии Staphylococcus aureus, содержащего большое количество поперечных связей. (Из работы: Ленинджер А., Основы биохимии, т. 1—3. — М.: Мир, 1985, т. 1, с. 317.)
На этом, однако, рассказ о структуре клеточных стенок бактерий не кончается. У многих видов микроорганизмов внешняя мембрана окружена капсулой или слизистым слоем, по химической природе представляющим собой полисахарид. Капсула одного из штаммов пневмококков (бактерий, вызывающих пневмонию) построена из чередующихся остатков глюкозы и глюкуроновой кислоты. Не имеющие такой полисахаридной капсулы мутанты не обладают патогенными свойствами. Полисахаридная капсула некоторых бактерий может растворяться в среде. Производство внеклеточных полисахаридов является важным промышленным процессом, который, однако, существенно осложняется неньютоновским характером течения жидкой среды (гл. 8). Слизистые слои, кроме того, принимают участие при флокуляции бактерий, являющейся важным этапом процессов обработки сточных вод методом активного ила.
Цитоплазматическая мембрана дрожжевых клеток состоит из липидов, белков и полисахаридов, содержащих остатки маннозы. Если двигаться дальше к внешней оболочке клетки, то за цитоплазматической мембраной будет расположено периплазматическое пространство, окруженное в свою очередь клеточной стенкой. В пекарских дрожжах, Saccharomyces cerevisiae, клеточная стенка содержит от 6 до 8% белков (в том числе несколько ферментов), а также приблизительно по 30% (по массе) глюкана (полисахарида, построенного из остатков d-глюкозы, соединенных ß-1,6-связями, а также поперечными ß-1,3-связями) и маннана (полиманнозы с а-1,6-связями и a-1,2-боковыми цепями). При обработке S. cerevisiae ферментом, гидролизующим глюкан, например 1,3-глюканазой, можно получать соответствующие протопласты. Такую обработку обычно проводят при введении рекомбинантных молекул ДНК в дрожжевые клетки.
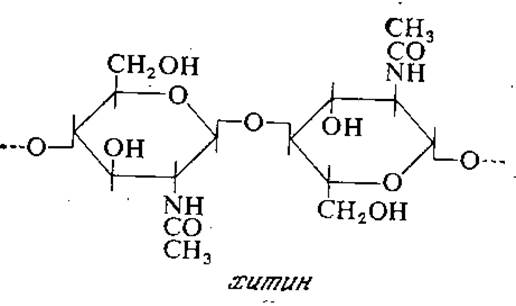
Клеточные стенки дрожжей и многих плесеней содержат хитин, макромолекула которого построена из остатков N-ацетилглюкозамина, соединенных гликозидными ß-1,4-связями (строение хитина показано на следующей странице).
Животные клетки, существующие обычно в строго контролируемом изотоническом окружении, не имеют клеточных стенок. Помимо фосфолипидов и белков в их плазматических мембранах содержится от 2 до 10% углеводов. Последние обнаружены на внешней поверхности всех клеток млекопитающих, изученных до настоящего времени; они связаны с липидами и белками в виде гликолипидов и гликопротеинов соответственно.