Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Стехиометрия и энергетика метаболических превращений
Фотосинтез и его связь с первичным источником энергии
Транспорт электронов и фотофосфорилирование
В фотосистемах I и II поглощение двух световых квантов (фотонов) хлорофиллом и другими пигментами приводит к возбуждению электрона в реакционном центре (например, Р700 в случае фотосистемы I; см. рис. 5.10). Обладающий достаточно высокой энергией электрон включается в цепь электронного транспорта. Затем электрон «падает» на низший энергетический уровень (этот процесс изображен на рис. 5.10 как повышение значения стандартного электродного потенциала Е⁰'). В фотосистеме I электрон в конечном счете используется для восстановления NADP+, а возбужденные электроны фотосистемы II передаются фотосистеме I, причем в процессе их транспорта каждая пара возбужденных электронов участвует в фосфорилировании одной молекулы ADP. Таким образом, общая стехиометрия реакций, индуцируемых фотонами в фотосинтезирующих эукариотах, выглядит следующим образом:
H2O + 4hv + NADP+ + ADP + Pi→
→ NADPH + H+ + 1/2O2 + (ATP + H2O) (5.28)
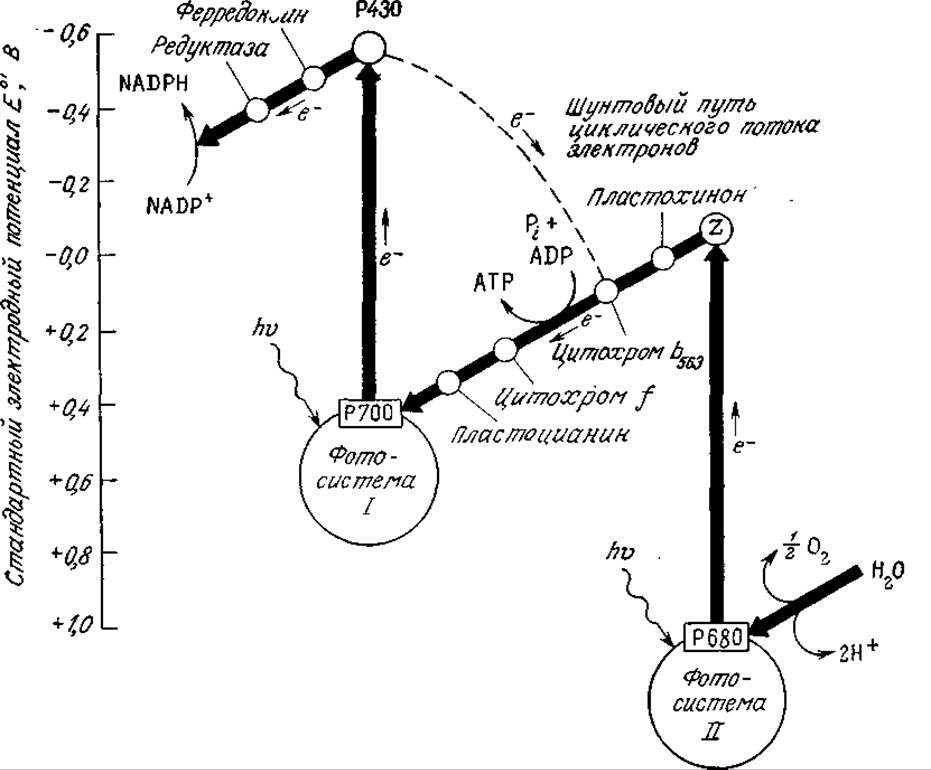
РИС. 5.10. Инициированное поглощением света возбуждение электронов (вертикальные стрелки) и их транспорт в фотосинтезирующих системах. (Ленинджер А., Основы биохимии, т. 1—3. — М.: Мир, 1985.)
В используемом диапазоне энергий светового возбуждения только при совместной работе двух фотосистем вода может служить донором (E⁰' = + 0,82 В), a NADP+ — акцептором (Е0' = -0,32 В) электронов. Уравнение (5.10) объясняет, почему, например, в бактериях, использующих в качестве донора электронов H2S, фотосинтез может осуществляться только фотосистемой I: это связано с величиной стандартного электродного потенциала восстановления серы Е⁰', равного —0,23 В.
Поглощенная световая энергия может участвовать в регенерации дополнительных количеств АТР по пути циклического фотофосфорилирования, изображенному на рис. 5.10 штриховой линией. В этом случае возбужденные электроны транспортируются от Р700 к Р430, затем к цитохрому b563 и снова к Р700; па последнем этапе этого пути электроны принимают участие в фосфорилировании ADP по следующему простому уравнению:
3hv + ADP + Pi → АТР + Н2O (5.29)
Использование накопленных рассмотренными выше путями энергии и восстанавливающих эквивалентов для синтеза глюкозы из СO2 и Н2O мы опишем в следующем разделе. Здесь же отметим, что изменение свободной энергии в реакции фотосинтеза
6СО2 + 6Н2О → C6H12O6 + 6O2 (5.30)
равно +686 ккал/моль. Поскольку в любой из фотосистем для возбуждения одного электрона необходимы два фотона и поскольку генерирование одной молекулы O2 сопровождается переносом четырех электронов, то образование 6O2 требует поглощения 48 квантов света. Согласно уравнению (5.27), энергия фотонов зависит от длины их волны. Если принять λ = 700 нм, то минимальное количество световой энергии, необходимой для инициирования реакции (5.30) с помощью фотосинтезирующей системы растений, составит 48∙41 = 1968 ккал/моль глюкозы. Следовательно, в первом приближении эффективность утилизации поглощенного света будет равна 686: 1968 = 0,35 = 35%. Действительную эффективность фотосинтеза в растениях и микроорганизмах мы рассмотрим позднее, в ходе обсуждения общей стехиометрии метаболизма.