Основы биохимии - Филиппович Ю. Б. 1999
Липиды и их обмен
Сложные липиды
Фосфолипиды. Фосфолипиды — сложные эфиры многоатомных спиртов с высшими жирными кислотами, содержащие остатки фосфорной кислоты и связанные с нею добавочные группировки (азотистые основания, аминокислоты, глицерин, инозит и др.).
Из многоатомных спиртов в составе различных фосфолипидов найдено три: глицерин, миоинозит и сфингозин:

В соответствии с этим фосфолипиды делят на три группы: глицерофосфолипиды, инозитфосфолипиды и сфингофосфолипиды. Глицерофосфолипиды часто называют фосфатидами, так как их можно рассматривать как производные фосфатидной кислоты (см. ниже), а инозитфосфолипиды — фосфоинозитидами.
В качестве высших жирных кислот в молекулах фосфолипидов содержатся пальмитиновая, стеариновая, линолевая, линоленовая и арахидоновая кислоты, а также лигноцериновая, нервоновая и др. (см. табл. 25).
В зависимости от типа фосфолипида в построении его молекулы принимают участие один или два остатка высшей жирной кислоты. Фосфорная же кислота входит, как правило, в состав фосфолипидов в количестве одной молекулы.
Лишь некоторые виды инозитфосфолипидов содержат два и более остатка фосфорной кислоты.
Азотсодержащие составляющие фосфолипидов разнообразны. Наиболее часто встречаются этаноламин, холин и серин (см. ниже). Из химического строения фосфолипидов ясно, что в их молекулах есть участки, способные диаметрально противоположно взаимодействовать с молекулами растворителя.
Углеводородный радикал остатка (или остатков) высших жирных кислот формирует лиофобную часть, а остатки фосфорной кислоты и азотистого основания, способные ионизироваться, — лиофильную. Благодаря этой особенности фосфолипиды, видимо, участвуют в обеспечении односторонней проницаемости мембран субклеточных структур.
Фосфолипиды — твердые вещества жироподобного вида; они бесцветны, но быстро темнеют на воздухе вследствие окисления по двойным связям входящих в их состав непредельных кислот. Хорошо растворяются в бензоле, петролейном эфире, хлороформе и т. п. Растворимость в спирте, ацетоне и серном эфире у разных групп фосфолипидов различна. В воде они не растворимы, но могут образовывать стойкие эмульсии, а в некоторых случаях — коллоидные растворы.
Фосфолипиды найдены в животных и растительных организмах, но особенно много содержит их нервная ткань человека и позвоночных животных. У беспозвоночных содержание фосфолипидов в нервной системе в 2—3 раза ниже. Много фосфолипидов в семенах растений, сердце и печени животных, яйцах птиц и т. п. Специфическими фосфолипидами обладают микроорганизмы.
Фосфолипиды легко образуют комплексы с белками и в виде фосфолипопротеинов присутствуют во всех клетках живых существ, участвуя главным образом в формировании клеточной оболочки и внутриклеточных мембран.
Глицерофосфолипиды, или фосфатиды, — сложные эфиры глицерина, высших жирных кислот, фосфорной кислоты и азотистого основания. Их рассматривают как производные фосфатидной кислоты, откуда и происходит само название этой группы фосфолипидов:
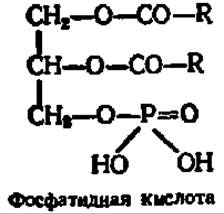
В зависимости от характера азотистого основания среди фосфатидов различают фосфатидилхолин (лецитины), фосфатидилколамин (кефалины), фосфатидилсерин и фосфатидилтреонин:
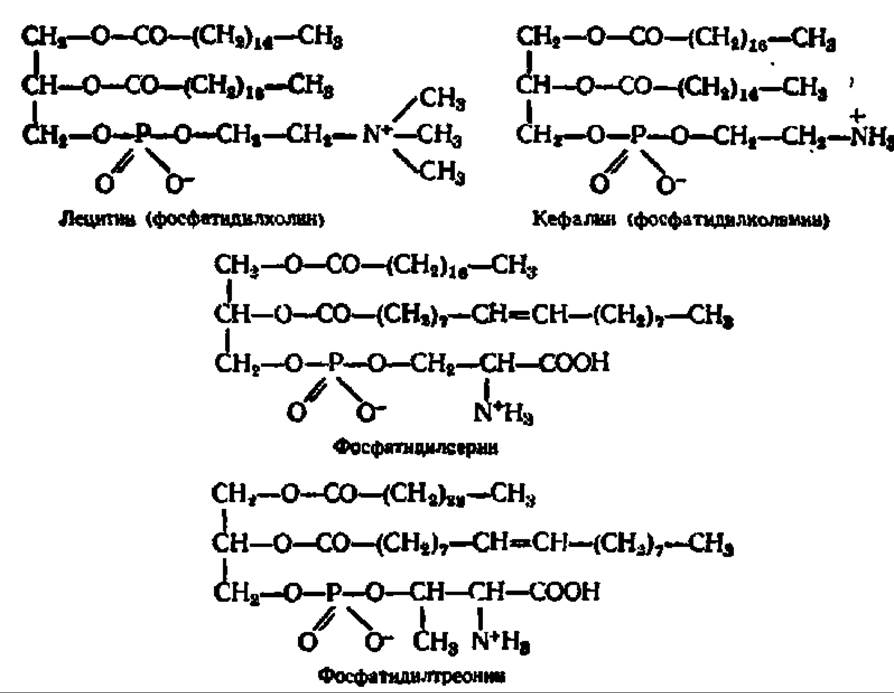
Из них лецитины наиболее распространены в природе. Наличие в лецитинах холина в качестве азотистого основания впервые было установлено К. С. Дьяконовым (1867).
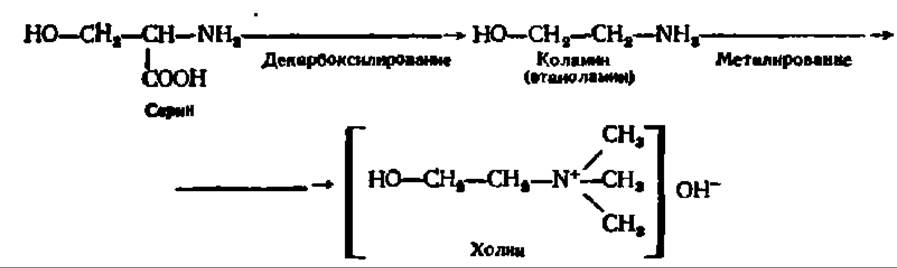
Три первых вида азотсодержащих фосфатидов, видимо, могут переходить друг в друга, так как они отличаются лишь строением азотистых оснований, между которыми возможна, например, такая генетическая связь:
Некоторые фосфатиды, открытые сравнительно недавно, не содержат азотистого основания, место которого в молекуле в этом случае занимают глицерин и его производные:
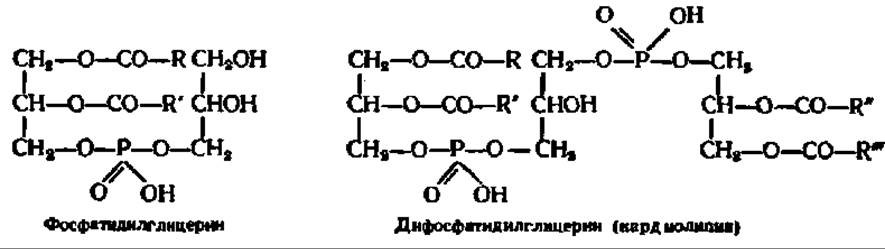
Фосфатидилглицерин является обязательной составной частью хлоропластов и в небольших количествах присутствует в бактериальных клетках и тканях животных. Кардиолипин — одно из существенно необходимых
соединений в составе митохондриальных мембран, особенно в митохондриях сердечной мышцы; он найден не только у животных, но и в растениях и у бактерий. И тот и другой способны аминоацилироваться по остатку глицерина с образованием аминоацилфосфатидил- и аминоацилдифосфатидил- глицеринов (их другое название — липоаминокислоты), создающих в хлоропластах, где они найдены (М. И. Молчанов, 1964), резерв аминокислот для биосинтеза белков.
Обладая асимметрическим строением (2-й углеродный атом остатка глицерина всегда асимметричен), фосфатиды оптически активны и образуют соответствующие стереоизомеры. Вместе с тем им свойственна изомерия за счет перестановки остатков высших жирных кислот из а- в ß-положение или наоборот.
Инозитфосфолипиды (фосфоинозитиды) представляют группу фосфолипидов, строение которой выяснено в последние 10—15 лет благодаря интенсивным работам по их химическому синтезу, осуществленным сотрудниками, принадлежащими к школам Н. А. Преображенского и Л. Д. Бергельсона. Простейший монофосфоинозитид построен так:
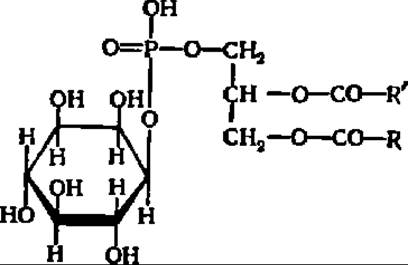
Как можно видеть, он похож на фосфатид, у которого азотистое основание заменено остатком инозита.
При наличии двух или трех остатков фосфорной кислоты в молекуле инозитфосфолипида все они связаны с остатком инозита. Инозитфосфолипиды такого типа выделены из мозга и получены синтетически. Особенно высоко их содержание в миелиновых оболочках нервных волокон спинного мозга. Скорость обмена фосфатных групп в фосфоинозитиддифосфатах намного превышает таковую в других фосфолипидах. Благодаря наличию диссоциированных фосфатных групп фосфоинозитиддифосфаты обеспечивают перенос ионов через биологические мембраны. Они же являются источником вторичных посредников, отщепляющихся в ответ на гормональный и иные сигналы (см с. 456 и 474). Многие фосфоинозитиды содержат углеводы, амины, аминокислоты и сфингозин.
Сфннгофосфолипиды в отличие от рассмотренных выше фосфатидов и инозитфосфолипидов содержат остаток высшей жирной кислоты, соединенный с двухатомным аминоспиртом (сфингозином) пептидной связью:
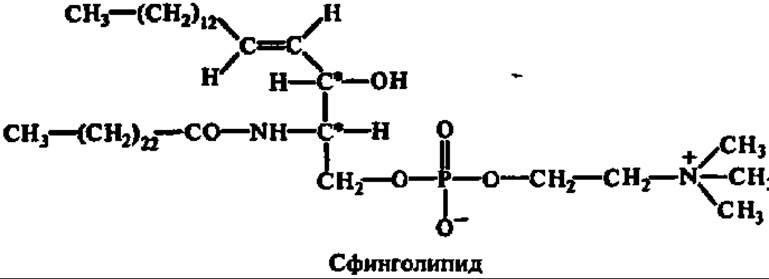
Овальные компоненты сфингофосфолипидов — фосфорная кислота и холин — присоединены так же, как у фосфатидов. Именно в сфинголипидах в значительных количествах обнаружены лигноцериновая и нервоиовая кислоты, менее характерные для остальных групп фосфолипидов.
Представители этой группы фосфолипидов, видимо, более характерны для животного, чем для растительного мира. Однако из фосфолипидов растительного происхождения (из кукурузного зерна) выделен аминоспирт, весьма похожий на сфингозин:
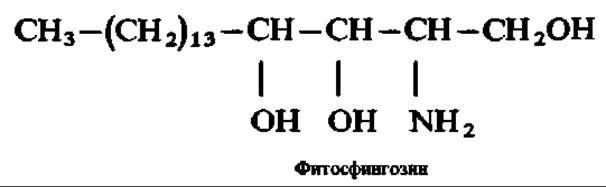
Этот же спирт найден в дрожжах и грибах, а недавно — в мозге и почках человека, что указывает на возможность существования аналогичных фосфолипидов в растительных и животных объектах.
Сфингофосфолипиды не растворимы в серном эфире, что используют при их отделении от фосфатидов. Они характеризуются также трудной растворимостью в ацетоне и большей устойчивостью к действию окислителей, чем фосфатиды.
Сфинголипидам свойственны весьма сложные пространственные конфигурации, связанные с возможностью оптической изомерии (два асимметричных углеродных атома в молекуле) и цис-транс-изомерии по месту двойной связи. Этим объясняется их органная и видовая специфичность. Кроме того, установлено, что органная специфичность сфинголипидов зависит от качественного состава высших жирных кислот: так, для сфинголипидов мозга характерно присутствие нервоновой кислоты.
Гликолипиды. Вторую группу сложных липидов образуют гликолипиды. Они характеризуются тем, что полярная моно- или олигосахаридная составляющая (глюкоза, галактоза, глюкозамин, галактозамин, их N-ацетильные производные и др.) через остаток многоатомного спирта (глицерин, сфингозин) соединяется с неполярными радикалами высших жирных кислот (пальмитиновой, стеариновой, олеиновой, лигноцериновой, нервоновой, цереброновой и др.) гликозидной и сложноэфирной связями.
Примерами являются моногалактозилглицерин и цереброн соответственно:
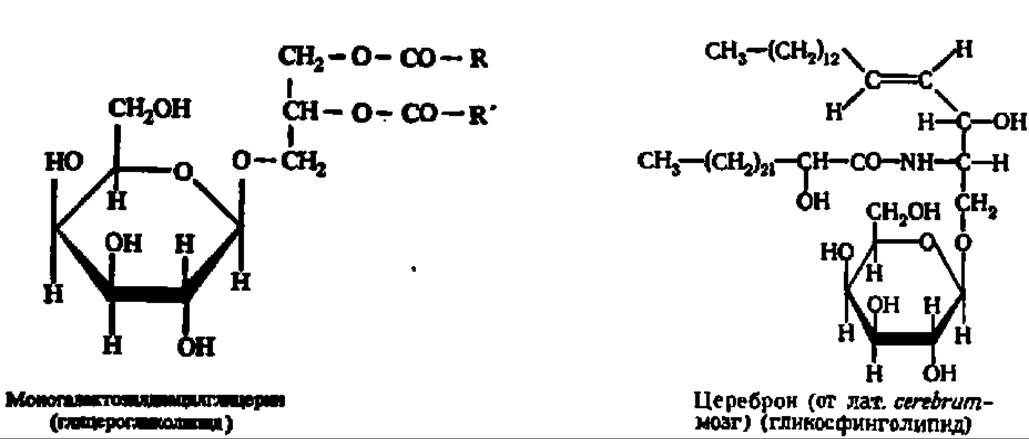
Глицерогликолипиды и гликосфинголипиды широко распространены в природе. Особое внимание сейчас привлекают гликосфинголипиды, содержащие олигосахаридную компоненту, например глобозиды:
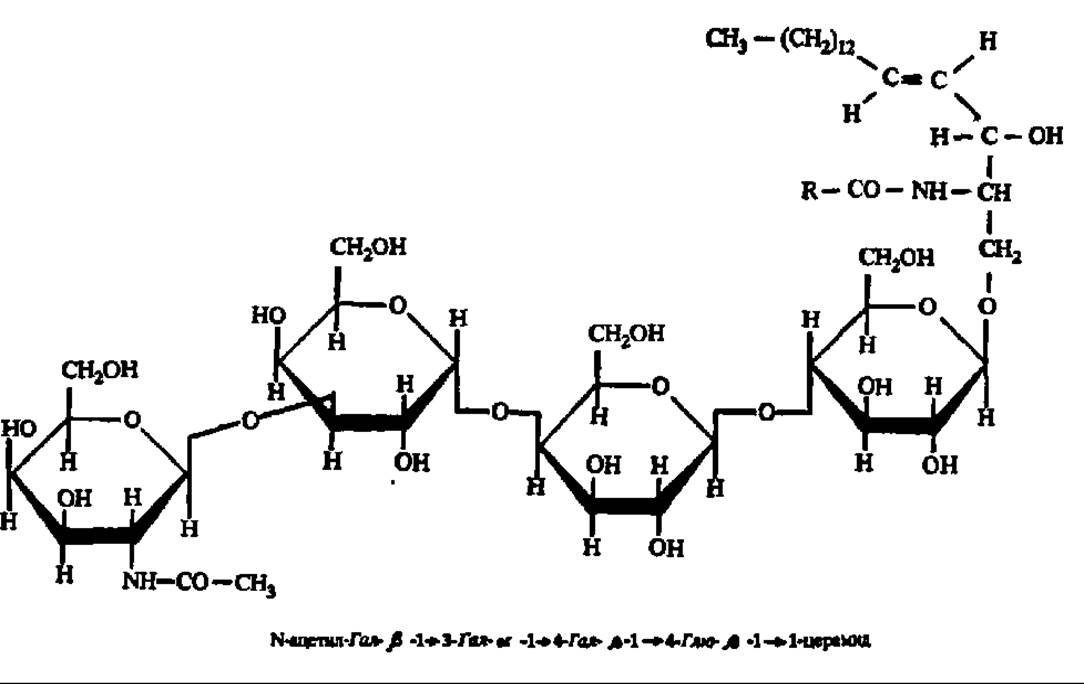
Уже изучено 110 глобо-, лакто- и ганглиосфинголипидов. Они регулируют рост клеток, являются маркерами трансформации нормальных клеток в раковые, обеспечивают компактизацию бластомеров во время дробления яйца, взаимодействуют с белковыми токсинами и выполняют ряд других важнейших функций.
В последнее время показано, что они участвуют в процессах рецепции гормонов, факторов роста, лейкинов, в регуляции роста и дифференцировки клеток, являются иммуномодуляторами и вторичными посредниками.
Диольные липиды. В природных липидах сложноэфирные связи с остатками высших жирных кислот и фосфорной кислотой, равно как и простые эфирные связи с остатками высших спиртов и углеводов, могут образовывать двухатомные спирты — этандиол, а также пропандиолы, бутандиолы и пентандиолы.
Такие липиды называют диольными липидами. Они широко распространены среди растений, животных и микроорганизмов в качестве минорных компонентов липидной фракции, хотя у морских беспозвоночных и рыб нередко составляют главную часть запасных липидов. Они же являются структурными элементами мембран эндоплазматической сети, но не митохондрий. Замечено, что они обмениваются энергичнее, чем липиды, где функцию спирта выполняет глицерин. Известно 9 групп диольных липидов, выделенных из различных объектов.
Орнитинолипиды. Первые указания о существовании этого нового вида липидов относятся к 1963 г. В течение двух последующих десятилетий структура многих из них была расшифрована и высказаны предложения об их функциональной роли.
Орнитинолипиды характерны для микроорганизмов. Их непременными составными частями являются высшие жирные ß-оксикислоты и аминокислота орнитин (или лизин); в зависимости от типа строения могут присутствовать также остатки этиленгликоля или 1;3-пропандиола. Наиболее распространенные в природе орнитинолипиды построены следующим образом:
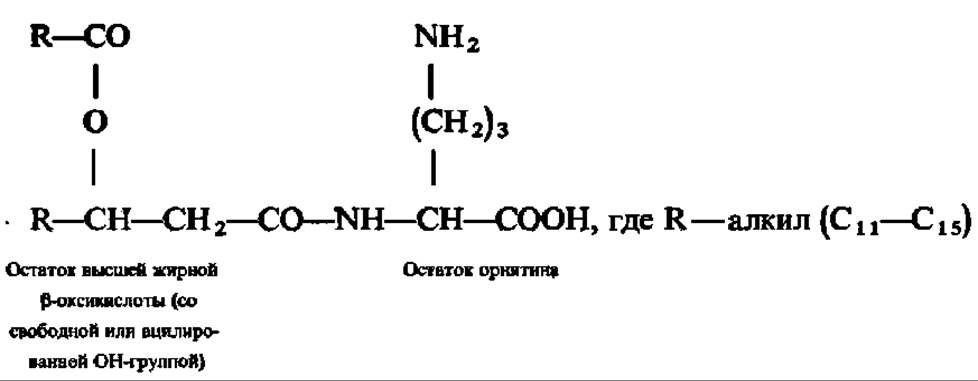
Установлено, что орнитинолипиды в микробной клетке способны заменять фосфатидилэтаноламин в качестве структурного элемента цитоплазматических мембран. Это особенно ярко проявляется в условиях фосфатного голодания.