ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 1. ОСНОВЫ БИОХИМИИ СТРОЕНИЕ И КАТАЛИЗ - 2011
ЧАСТЬ I. СТРОЕНИЕ И КАТАЛИЗ
4. ТРЕХМЕРНАЯ СТРУКТУРА БЕЛКОВ
4.3. Третичная и четвертичная структуры белка
Архитектура белка — введение в третичную структуру.Общая организация всех атомов белка в пространстве называется третичной структурой. Если термин «вторичная структура» относится к пространственной организации аминокислотных остатков, расположенных близко друг от друга в полипептидной цепи, третичная структура подразумевает способ организации молекулы белка в целом. Аминокислотные остатки, находящиеся далеко друг от друга в полипептидной цепи и участвующие в образовании различных типов вторичной структуры, могут взаимодействовать друг с другом в полностью свернутом белке. Локализация изгибов цепи (в том числе и β-поворотов), а также направление и угол поворота определяются числом и расположением специфических остатков, таких как Pro, Thr, Ser и Gly. Взаимодействующие между собой участки полипептидной цепи удерживаются в характерном положении в пространстве в результате действия слабых сил (а иногда и с помощью ковалентных связей, таких как дисульфидные мостики).
Некоторые белки состоят из двух или большего чиста отдельных полипептидных цепей, называемых субъединицами, которые могут быть как одинаковыми, так и разными. Взаимное расположение субъединиц белка в пространстве определяет четвертичную структуру белка.
При рассмотрении этих более высоких уровней организации белковой структуры удобно разделить все белки на два основных класса: фибриллярные белки, полипептидные цепи которых организованы в виде длинных нитей или волокон, и глобулярные белки, чьи полипептидные цепи образуют сферические структуры — глобулы. Эти два класса белков заметно различаются по структуре: фибриллярные белки обычно характеризуются одним типом вторичной структуры, а отдельные участки глобулярных белков часто имеют разную вторичную структуру. Два класса белков различаются и по основным функциям: те структуры в организмах позвоночных, которые определяют их жесткость, форму и защиту от внешних воздействий, построены из фибриллярных белков, а большинство ферментов и регуляторных белков имеют глобулярное строение. Несколько фибриллярных белков сыграли очень важную роль в развитии современных представлений о структуре белка и служат прекрасными примерами, иллюстрирующими взаимосвязь между структурой белка и его функцией. Мы начнем с изучения третичной структуры фибриллярных белков, а затем обратимся к более сложному типу укладки полипептидной цепи, характерной для глобулярных белков.
Фибриллярные белки адаптированы для выполнения структурной функции
Архитектура белка — третичная структура фибриллярных белков.На примере α-кератина, коллагена и фиброина шелка можно прекрасно продемонстрировать связь между структурой белка и его функцией (табл. 4-2). Фибриллярные белки обладают свойствами, определяющими прочность и (или) гибкость тех структур, в построении которых они принимают участие. В любом случае основной структурной единицей таких белков является повторяющийся элемент вторичной структуры. Фибриллярные белки не растворяются в воде, что связано с высокой концентрацией гидрофобных аминокислотных остатков как внутри белка, так и на его поверхности. Гидрофобные поверхности белков в значительной степени скрыты, поскольку сходные полипептидные цепи укладываются рядом и образуют надмолекулярные комплексы. Простота строения фибриллярных белков делает их прекрасным примером для изучения некоторых основных принципов организации белка, которые мы обсуждали выше.
Таблица 4-2. Вторичная структура и свойства фибриллярных белков
Структура |
Характеристики |
Примеры |
α-Спирали, сшитые дисульфидными связями |
Прочные, нерастворимые в воде защитные структуры различном жесткости |
α-Кератин волос, перьев и ногтей |
β-Конформация |
Мягкие, гибкие волокна |
Фиброин шелка |
Тройная спираль коллагена |
Высокая прочность и эластичность при растяжении |
Коллаген в сухожилиях и в костной ткани |
α-Кератин. α-Кератины созданы природой для придания жесткости структурам организма. У млекопитающих эти белки — основные компоненты волос, шерсти, ногтей, когтей, перьев, рогов, копыт и многих других поверхностных образований кожи. α-Кератины являются представителями более широкого семейства белков — промежуточных филаментов. Другие белки промежуточных филаментов обнаружены в цитоскелете клеток животных. Все белки этого семейства выполняют структурную функцию и имеют такие же структурные особенности, как α-кератины.
Молекула α-кератина представляет собой правую спираль, которая встречается во многих других белках. Фрэнсис Крик и Лайнус Полинг в начале 1950-х гг. независимо друг от друга пришли к выводу, что α-спираль кератина организована в виде двухспиральной катушки. Две ориентированные параллельно нити α-кератина (т. е. с N-концами с одной и той же стороны) закручиваются одна вокруг другой, образуя суперскрученную спираль. Такая суперскрученность придает всей структуре дополнительную прочность — точно так же, как скрученные веревки придают прочность канату (рис. 4-10). Искривление оси α-спирали, образующей спиральную катушку, объясняет расхождение между расчетным значением витка спирали (5,4 Â), полученным Полингом и Кори, и экспериментальным значением (5,15-5,20 А), полученным при рентгеноструктурном анализе волоса (с. 177). Супервитки α-спирали образуют левую спираль, т. е. направлены противоположно направлению витков α-спирали. Поверхности соприкосновения двух α-спиралей состоят из гидрофобных аминокислотных остатков, а их R-группы сцеплены друг с другом регулярным образом. Это создает компактную упаковку полипентидных цепей в левозакрученной суперспирали. Неудивительно, что α-кератины богаты такими гидрофобными остатками, как Ala, Val, Leu, Ile, Met и Phе.
Рис. 4-10. Структура волоса, а) α-Кератин волос образован вытянутыми α-спиралями с утолщениями на N- и С-концах аминокислотных цепей. Спирали попарно оборачиваются одна вокруг другой, в результате чего образуется левозакрученная двойная суперспираль. Эти суперспирали в свою очередь организованы в другие высокоупорядоченные структуры — протофиламенты и протофибриллы. Около четырех протофибрилл (32 цепи α-кератина, собранные вместе) образуют промежуточный филамент. В других структурах также наблюдается сцепление отдельных суперспиралей, но направление образующихся спиралей и структурные детали неизвестны, б) Волос представляет собой пучок из множества нитей α-кератина, построенных таким образом, как показано на рисунке (а).
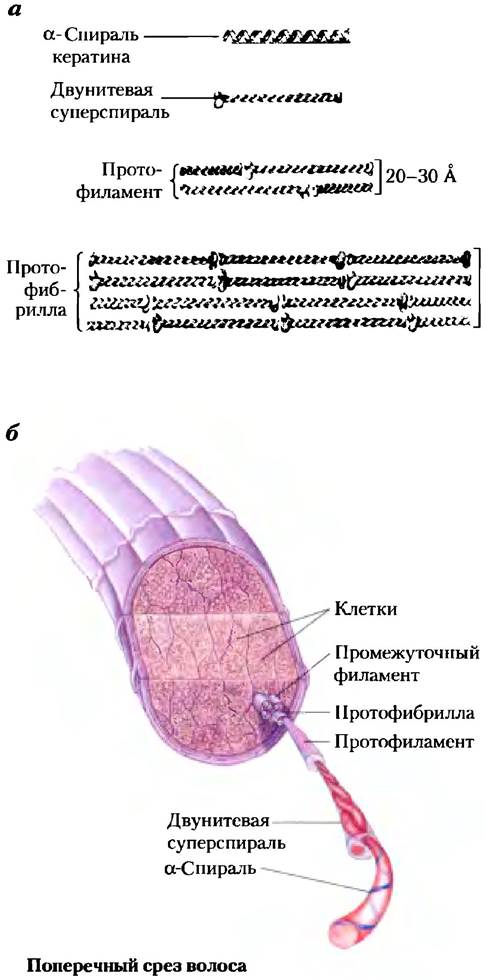
Отдельные полипептидные цепи в двойной спирали α-кератина имеют сравнительно простую третичную структуру, в которой доминируютα-спиральные участки, сами образующие левые суперспирали вокруг своей оси. Взаимодействие между отдельными α-спиралями представляет собой пример четвертичной структуры. Спиральные катушки такого типа являются распространенным структурным элементом в фибриллярных белках и в мышечном белке миозине (рис. 5-27). Четвертичная структура α-кератинов может быть и сложнее. Множество спиральных катушек могут быть собраны в большой надмолекулярный комплекс; такую структуру, например, имеют молекулы α-кератина в промежуточных филаментах волос (рис. 4-10, б).
Прочность фибриллярных белков дополнительно повышается за счет ковалентных сшивок между отдельными полипептидными цепями и между отдельными нитями в надмолекулярной структуре. В α-кератинах в качестве таких стабилизирующих четвертичную структуру связей выступают дисульфидные мостики (доп. 4-2). В самых жестких и прочных α-кератинах, таких как в роге носорога, до 18% аминокислотных остатков приходится на долю цистеина, участвующего в образовании дисульфидных связей.
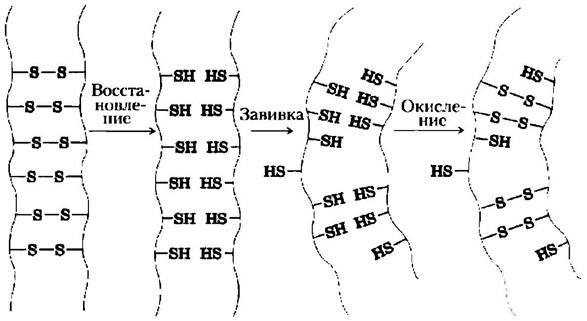
Перманентная завивка волос — пример биохимический технологии
Известно, что влажные волосы при нагревании способны растягиваться. На молекулярном уровне это означает, что α-спирали α-кератина растягиваются до тех пор, пока не примут плоскую β-конформацию. При охлаждении они самопроизвольно возвращаются в конформацию α-спирали. Такая характерная эластичность α-кератинов и наличие множественных дисульфидных мостиков являются молекулярной основой химической завивки волос. Волосы сначала накручивают на бигуди, чтобы получились кудри определенной формы, а затем при нагревании обрабатывают раствором восстанавливающего агента, которым обычно служит вещество с тиоловой или сульфгидрильной группой (-SН). Восстановитель расщепляет дисульфидные связи, в результате чего из каждой связи образуются два остатка цистеина. Влажность и тепло разрушают водородные связи и заставляют раскручиваться α-спирали. Через некоторое время восстановитель смывают и добавляют раствор окислителя, под действием которого образуются новые дисульфидные связи между парами остатков цистеина, расположенных на соседних полипептидных цепях (но это совсем
не те пары остатков Суs, что были раньше). После того как волосы высохли и охладились, полипептидные цени вновь принимают конформацию α-спирали. Волосы вьются, поскольку новые дисульфидные связи зафиксировали пучки α-спиралей кератиновых волокон в определенном положении. Перманентная завивка или выпрямление волос на самом деле держатся не бесконечно, поскольку волосы отрастают. Старые волосы постепенно заменяются новыми с естественным расположением дисульфидных связей.
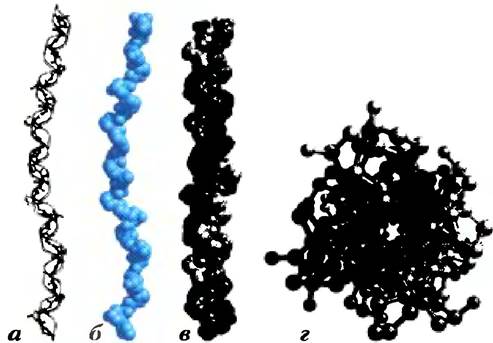
Коллаген. Подобно α-кератинам, коллаген обеспечивает прочность структур организма. Он присутствует в соединительных тканях, таких как сухожилия, хрящи, органическое вещество костного матрикса и роговица глаза. Спираль коллагена — это уникальная вторичная структура, отличающаяся от α-спирали. Полипептидные цепи коллагена уложены в левую спираль, один виток которой построен из трех аминокислотных остатков (рис. 4-11). Эти спирали также образуют суперспиральные структуры, однако имеющие совсем другое строение: три полипептидных цепи, называемые α-цепями (не путайте с α-спиралями), закручены одна вокруг другой (рис. 4-11, в). В коллагене суперспираль закручена вправо — в противоположном направлении, чем левые спирали α-цепей.
Рис. 4-11. Структура коллагена (по данным РDВ ID 1СGD). а) α-Цепьколлагенаимеетуникальную вторичную структуру. Повторяющаяся трипептидная последовательность Glу-Х-Рго или Glу-Х-4-Нур образует левую спираль с тремя остатками в одном витке. В изображенной модели использована последовательность Glу-Рго-4-Нур. б) Пространственная модель той же α-цепи. в) Три подобные цепи (изображены серым, синим и лиловым цветом) обвивают друг друга, образуя правую спираль. г) Шаро-стержневая модель тройной суперспирали коллагена (вид с одного конца). Красным цветом показаны остатки Glу. Только глицин, имеющий самый маленький размер, может поместиться в таком узком пространстве, как место контакта трех полипептидных цепей. Размер шариков в данной модели не соответствует ван-дер- ваальсовым радиусам атомов. Внутри этой тройной суперспирали нет пустого пространства, как изображено на рисунке; на самом деле полипептидные цепи находятся в очень тесном контакте.
У позвоночных существует несколько типов коллагенов. Обычно эти белки содержат около 35% Gly, 11% Ala, а также 21% Pro и 4-Нур (аминокислота 4-гидроксииролин, рис. 3-8, а). Из коллагена получают пищевой желатин; он имеет небольшую пищевую ценность в качестве белка, поскольку содержит чрезвычайно мало аминокислот, важных для питания человека. Необычный аминокислотный состав коллагена определяет его уникальную структуру. Последовательность аминокислот в коллагене обычно представляет собой чередование трипептидных звеньев с формулой Gly-X-Y, где часто X — пролин Pro, a Y — 4-гидроксипролин Нур. Только остаток Gly может поместиться в узкую щель между отдельными a-цепями (рис. 4-11, г), а остатки Pro и 4-Нур обеспечивают крутой поворот спирали коллагена. Аминокислотная последовательность коллагена и наличие суперспирали приводят к тому, что три полипептидные цепи очень плотно упакованы. В структуре коллагена 4-гидроксипролин очень важен; эта аминокислота сыграла определенную роль в истории человечества (доп. 4-3).
Тесное сплетение a-цепей в тройной спирали коллагена в пересчете на единицу площади поперечного сечения обеспечивает при растяжении более высокую прочность, чем прочность стальной проволоки. Нити коллагена (рис. 4-12) представляют собой надмолекулярные структуры, состоящие из тройных спиралей коллагена (иногда называемых тропоколлагеном), связанных между собой различным образом, что приводит к образованию структур с разной прочностью. Связывание a-цепей коллагена между собой происходит необычно — посредством ковалентной связи с участием остатков Lys, HyLys (5-гидроксилизина, рис. 3-8. а) или His, время от времени встречающихся в позициях X или Y трипептидного звена коллагена. Подобное связывание приводит к образованию необычных аминокислотных остатков, таких как дегидрогидроксилизинонорлейцин. По мере старения организма в фибриллах коллагена появляется больше поперечных сшивок, что делает соединительные ткани все более жесткими и хрупкими.
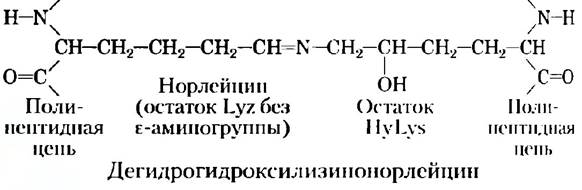
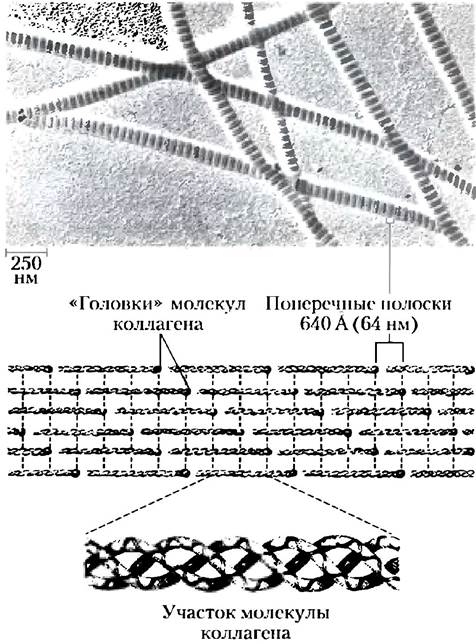
Рис. 4-12. Структура фибрилл коллагена. Молекула коллагена (Мr = 300 000) имеет форму палочки длиной около 3000 А и шириной лишь 15 А. Три закрученные в спираль α-цепи коллагена могут иметь различные последовательности, но каждая состоит примерно из 1000 аминокислотных остатков. Фибриллы коллагена состоят из вытянутых молекул коллагена, сшитых между собой для дополнительной прочности. Специфическое расположение и степень сшивки в разных тканях различаются. На электронных микрофотографиях фибриллы коллагена имеют характерные поперечные полосы. «Головки» молекул коллагена расположены через каждые 640 А.
Дополнение 4-3. МЕДИЦИНА. Почему морякам, путешественникам и студентам нужно есть свежие фрукты и овощи
...после этого несчастья, а также из-за общей нездоровой обстановки в этом краю, где никогда не выпадает ни капли дождя, нас всех поразила «лагерная болезнь», заключавшаяся в том, что наши конечности стали усыхать, кожа на ногах, подобно старому башмаку, стала покрываться черными пятнами, как будто плесенью, опухали десны, и никто из заболевших не вырвался из лап смерти. Знак был такой: если пошла из носа кровь, то смерть близко...
Из воспоминаний лорда Джоинвилля, около 1300 г.
Данный фрагмент текста описывает состояние армии Людовика IX к моменту окончания седьмого крестового похода (1248-1254), незадолго до битвы при Фа- раскуре, где ослабленная цингой армия крестоносцев была окончательно разбита египтянами. В чем была причина болезни, поразившей воинов в XIII в.?
Цинга возникает при недостатке витамина С (аскорбиновой кислоты). Витамин С, кроме прочего, необходим для гидроксилирования пролина и лизина в составе коллагена; при дефиците витамина С происходит общее поражение соединительной ткани. Проявлениями прогрессирующего заболевания являются частые небольшие кровотечения, связанные с хрупкостью сосудов, выпадение зубов, медленное заживление ран и открытие старых ран, боль и слабость костей, а в конечном итоге - остановка сердца. Отмечается подавленное настроение и повышенная чувствительность к раздражителям разного рода. При недостаточности витамина С средней тяжести возможны утомляемость, раздражительность и более тяжелое протекание инфекционных заболеваний дыхательных путей. Многое животные синтезируют витамин С в больших количествах путем четырехстадийного ферментативного превращения глюкозы в аскорбат. Однако в процессе эволюции человек и некоторые другое животные (гориллы, морские свинки и фруктовые летучие мыши) утратили последний из четырех ферментов, необходимых для осуществления этого процесса, и поэтому должны получать аскорбиновую кислоту с пищей. Витамин С содержится во многих фруктах и овощах. Однако до 1800 г. он часто отсутствовал в продуктах, запасаемых на зиму или для дальних путешествий.
Цинга была описана египтянами в 1500 г. до н. э., а затем Гиппократом в V в. до н. э. Хотя цинга оказывала немаловажное влияние на ход средневековых войн и периодически давала о себе знать в зимнее время в странах с холодным климатом, более пристальное внимание к этому заболеванию было обращено только во времена великих географических открытий — с 1500 по 1800 г., которые потребовали дальних путешествий. В первом кругосветном путешествии, совершенном Фернаном Магелланом (1520 г.), цинга унесла жизнь более 80% членов экспедиции. Васко да Гама по той же причине потерял две трети своей команды в первой экспедиции в Индию (1499 г.). Во время второго путешествия Жака Картье на реку Святого Лаврентия его экспедиция испытала множество превратностей судьбы, ей грозила полная гибель; благо, что местные жители научили команду заваривать кедровый чай, который излечивал и предотвращал цингу (поскольку содержал витамин С). По оценкам, в период от 1600 до 1800 г. около миллиона моряков умерли от цинги. Зимние вспышки цинги в Европе постепенно исчезли в XIX в., что, по-видимому, было связано с распространением картофеля, ввезенного из Южной Америки.
В 1747 г. шотландский врач Джеймс Линд, служивший в Королевской армии, провел первые в истории зарегистрированные контролируемые испытания. При длительном плавании на 50-пушечном военном корабле Сэлсбери он разделил 12 страдавших от цинги моряков на группы по два человека. Все 12 ели одну и ту же пищу, но получали разные лекарства, применявшиеся в то время для лечения цинги. Тс матросы, которые ели лимоны и апельсины, поправлялись и вновь были способны работать, те, кто пил кипяченый яблочный сок, поправлялись слабо, а состояние остальных ухудшалось. В 1753 г. Линд издал «Трактат о цинге», однако еще на протяжении 40 лет в Королевском флоте не предпринимались никакие меры. Наконец, в 1795 г. Британское адмиралтейство издало указ о введении концентрированного сока лимона или лайма в рацион всех британских моряков (отсюда пошло прозвище английских моряков «лайми»), Цинга оставалась бедствием в некоторых частях света до 1932 г., когда венгерский ученый Альберт Сент-Дьердьи, а также У. A. Boy и С. Г. Кинг из Университета Питтсбурга выделили и синтезировали аскорбиновую кислоту.
Джеймс Линд (1716-1794), военно-морской врач с 1739 по 1748 г.

L-Аскорбиновая кислота (витамин С) представляет собой белый кристаллический порошок, не имеющий запаха. Он хорошо растворяется в воде и почти не растворяется в органических растворителях. В виде сухого порошка и вне досягаемости прямых солнечных лучей вещество довольно стабильно. До сих нор нет единого мнения о суточной норме потребления витамина С. В США рекомендуемая суточная норма составляет 60 мг, в Австралии и Великобритании — 30-40 мг, в России — 100 мг. Иногда советуют принимать более высокие дозы витамина С, хотя до сих пор однозначно не установлено, приносит ли это дополнительную пользу. Интересно, что такое содержание витамина С, как в организме животных, способных его самостоятельно синтезировать, в организме человека достигается только при потреблении дозы, в сотни раз превосходящей рекомендованную суточную дозу. Кроме цитрусовых и многих других свежих фруктов хорошим источником витамина также являются перец, томаты, картофель и брокколи. При длительном приготовлении или хранении фруктов и овощей витамин С в них разрушается.
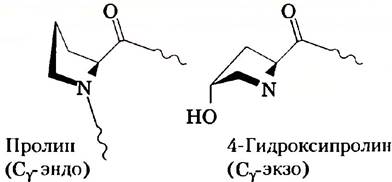
Итак, почему же витамин С так необходим для здоровья? Для нас особый интерес представляет его участие в формировании коллагена. Коллаген построен из повторяющихся трипептидных звеньев состава Gly-X-Y, где обычно X и Y — это Pro или 4-Нур, производное пролина 4(R)-L-гидроксипролин, который играет важную роль в фолдинге и поддержании структуры коллагена. Синтетические пептиды из 10 повторов Gly-Pro-Pro могут складываться в тройную спираль коллагена, однако эта структура плавится при 41 °С. Если же пептид состоит из звеньев состава Gly-Pro-Hyp, то точка плавления тройной спирали повышается до 69 °С. Стабильность коллагена напрямую связана с его тройной спиралью, строение которой независимо определили Хелен Берман и Адриана Загари с коллегами. Пролин обычно присутствует в двух конформациях, называемых Сγ-эндо и Сγ-экзо (рис. 1). Для образования спирали коллагена необходимо, чтобы остаток пролина в положении Y находился в Сγ-экзо-конформации, образованию которой способствует наличие гидроксильной группы у атома С-4 в 4-гидроксипролине. Структура коллагена также требует, чтобы остаток Pro в позиции X находился в конформации Сγ-эндо, и введение в эту позицию 4-Нур дестабилизирует спираль. Невозможность гидроксилировать Pro в позиции Y при дефиците витамина С приводит к нестабильности структуры коллагена и тем проблемам с соединительной тканью, которые возникают при цинге.
Рис. 1. Cγ-эндо-конформация пролина и Сγ-экзо-конформация 4-гидроксипролина.
Для гидроксилирования остатков Pro в предшественнике коллагена проколлагене необходим фермент пролил-4-гидроксилаза. У всех позвоночных этот фермент представляет собой α2β2-тетрамср с молекулярной массой 240 000. Активность пролилгидроксилазы связана с α-субъединицей. (Ученые были удивлены, обнаружив, что β-субъединины этого белка такие же, как в ферменте дисульфидизомеразе белков; эти субъединицы не участвуют в проявлении пролилгидроксилазной активности.) Каждая α-субъединица содержит один атом негемового железа (Fе2+); фермент относится к классу гидроксилаз, которым для проявления активности необходим α-кетоглутарат.
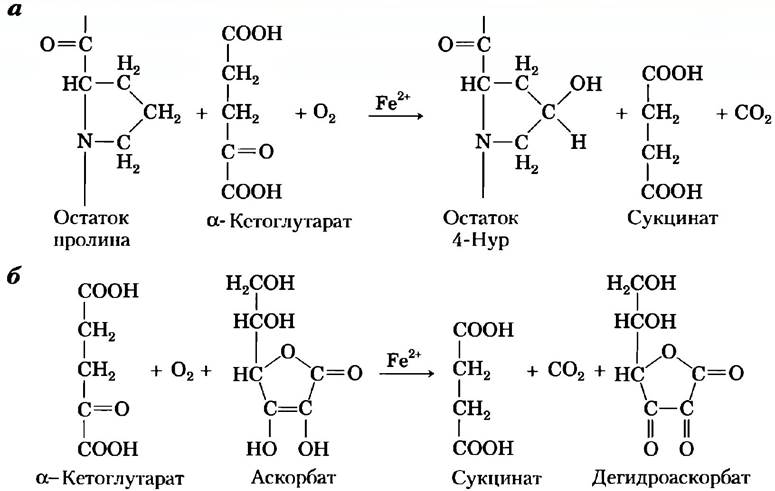
В норме в реакции с участием пролилгидроксилазы (рис. 2, а) с ферментом связана одна молекула кислорода и одна молекула α-кетоглутарата. α-Кетоглутарат подвергается окислительному декарбоксилированию с образованием СО2 и сукцината. Оставшийся на ферменте атом кислорода затем используется для гидроксилирования остатка Pro в проколлагене. Для этой реакции аскорбиновая кислота не нужна. Однако пролилгидроксилаза катализирует также окислительное декарбоксилирование α-кетоглутарата, не связанное с гидроксилированием пролина (рис. 2, б). В данной реакции происходит окисление гемового железа, а окисленная форма фермента не проявляет активности, т. е. не гидроксилирует пролин. Считается, что аскорбат в данной реакции служит для восстановления гемового железа и реактивации фермента.
Рис. 2. Реакции, катализируемые пролил-4-гидроксилазой. а) Нормальная реакция, сопряженная с гидроксилированием пролина: для этой реакции аскорбиновая кислота не нужна. Атомы кислорода, происходящие из молекулы О2, выделены красным цветом. б) Несопряженная реакция, в которой α-кетоглутарат подвергается окислительному декарбоксилированию, а гидроксилирования пролина не происходит. Аскорбиновая кислота потребляется в соответствии со стехиометрией уравнения реакции и превращается в дегидроаскорбат.
Люди болеют цингой до сих пор, причем не только в тех регионах, где существуют проблемы с продуктами питания, но и, как это не удивительно, в американских колледжах. Готовые салаты — единственные овощное блюдо в рационе питания некоторых студентов, да и фруктов они едят крайне мало. Проведенное в 1998 г. обследование 230 студентов Университета Аризоны показало, что 10% из них страдали серьезной недостаточностью витамина С, а у двух студентов уровень витамина был настолько низким, что они, по всей видимости, были больны цингой. Только половина студентов потребляли рекомендованную суточную норму витамина С.
Ешьте больше фруктов и овощей!
Обычно у млекопитающих присутствует более 30 структурных разновидностей коллагена, характерных для определенных видов тканей и несколько различающихся по аминокислотной последовательности и функциям. Некоторые генетические дефекты человека, связанные со структурой коллагена, служат примером тесной связи между аминокислотной последовательностью и трехмерной структурой этого белка. Несовершенный остеогенез характеризуется аномальным развитием костей у младенцев, а синдром Элерса-Данлоса связан с гипермобильностью суставов. Оба этих состояния могут быть летальными, и оба связаны с заменой Gly в каждой a-цепи на аминокислоту, имеющую более объемную R-группу, например, Cys или Ser (в двух названных заболеваниях происходит замена разных остатков Gly). Замена всего лишь одного аминокислотного остатка приводит к катастрофическим последствиям, поскольку нарушает трипептидное звено Gly-X-Y — основу уникальной спиральной структуры коллагена. Если учесть роль Gly в образовании тройной спирали коллагена (рис. 4-11, г), становится понятно, что его замена на любую другую аминокислоту обязательно приведет к нарушению структуры этого белка. ■
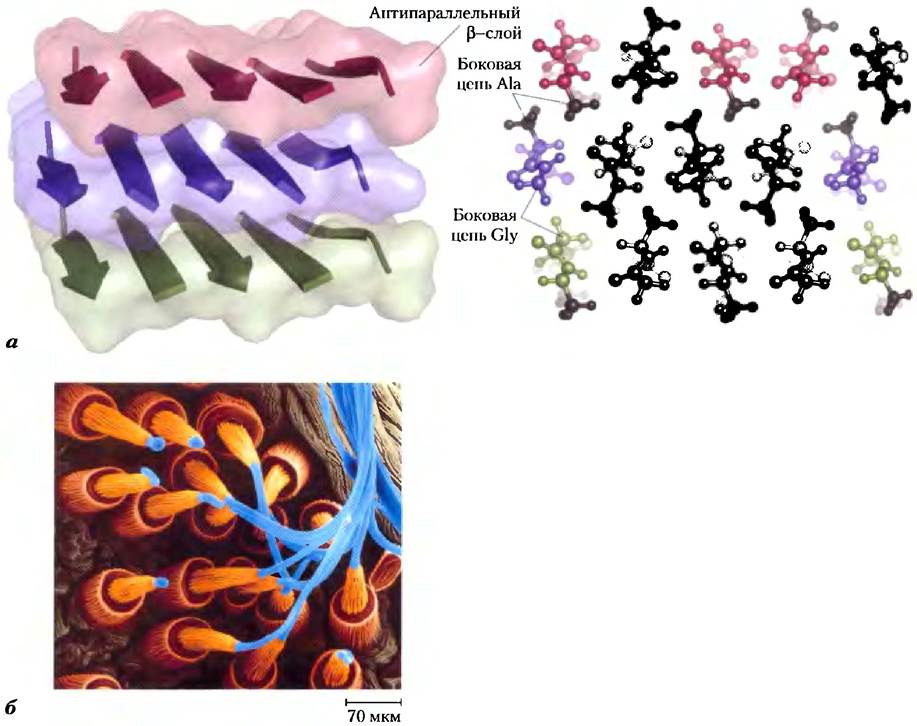
Фиброин шелка. Белок шелка фиброин синтезируется насекомыми и пауками. Его полипептидные цепи существуют преимущественно в p-конформации. Фиброин содержит много остатков Ala и Gly, что способствует плотной упаковке β-слоев и взаимному проникновению их R-групп (рис. 4-13). Вся структура в целом стабилизируется за счет протяженных водородных связей, образующихся между пептидными группами всех полипептидов в каждом β-слое, а также за счет оптимизации ван-дер-ваальсовых взаимодействий между слоями. Шелк не тянется, поскольку полипептидные цепи β-конформации и так максимально растянут (рис. 4-6). Однако подобная структура является довольно гибкой, поскольку слои удерживаются вместе не ковалентными связями, такими как дисульфидные мостики в α-кератинах, а множеством слабых взаимодействий.
Рис. 4-13. Структура шелка. И нити при изготовлении шелковых тканей, и паутина состоят из белка фиброина. а) Фиброин построен из уложенных в лист антипараллельных β-слоев. Полипептидные последовательности белка богаты остатками Ala (лиловые) и Gly (желтые Их небольшие R-группы способствуют плотной упаковке β-слоев, как показано на рисунке (вид сбоку). б) Окрашенная электронная микрофотография демонстрирует выход нитей фиброина (синего цвета) из прядильного органа паука.
Разнообразие структуры отражает функциональное многообразие глобулярных белков
В глобулярных белках отдельные участки полипептидной цепи (или несколько полипептидных цепей) свернуты между собой, образуя более компактную структуру, чем фибриллярные белки (рис. 4-14). Сворачивание (фолдинг) белка обеспечивает также структурное многообразие, необходимое белкам для осуществления самых разных биологических функций. Глобулярную структуру имеют ферменты, транспортные белки, двигательные белки, регуляторные белки, иммуноглобулины и множество других белков с разными функциями.
Рис. 4-14. Структуры глобулярных белков компактны и весьма разнообразны. Альбумин сыворотки человека (Мr = 64 500) имеет одну полипептидную цепь, состоящую из 585 аминокислотных остатков. На данной схеме сравниваются гипотетические размеры полипептидной цепи альбумина, находящейся исключительно в β-конформации или в виде α-спирали. Для сравнения также показан относительный размер белка в его нативной глобулярной форме, определенной методом рентгеноструктурного анализа. Чтобы достичь таких крохотных размеров, полипептидная цепь должна быть очень плотно упакована.

Число известных трехмерных структур белков исчисляется тысячами, и каждые два года, это число увеличивается еще в два с лишним раза. Такое богатство информации коренным образом меняет наши представления о структуре белка, о связи его структуры и функции, а также о тех эволюционных путях, которые привели к возникновению белков в их современном состоянии. Огромное многообразие белковых структур может показаться устрашающим. Однако по мере определения структур все новых и новых белков становится ясно, что все они относятся к ограниченному набору узнаваемых и устойчивых типов.
Наше обсуждение структур глобулярных белков мы начнем с рассказа о принципах строения, выведенных на основании структур самых первых изученных белков. Далее мы подробно остановимся на описании и сравнительной классификации надмолекулярных структур белков. Подобное обсуждение стало возможным только
благодаря такому мощному источнику информации, каким является Интернет, в частности благодаря находящимся в свободном доступе базам данных, таким как Protein DataBank (доп. 4-4).
Дополнение 4-4. Protein Data Bank
Количество известных трехмерных структур белков на сегодняшний день исчисляется тысячами и через каждые два года возрастает более чем вдвое. Это обилие информации произвело революцию в нашем понимании структуры белка, взаимосвязей их структуры и функции, а также путей эволюции белков к нынешнему состоянию, которые можно проследить по «фамильному» сходству, что приоткрывается нам по мере отбора и сортировки данных в базах данных. Один из важнейших для биохимиков доступных ресурсов — Protein Data Bank (PDB; www.rcsb.org).
PBD представляет собой архив экспериментально полученных трехмерных структур биологических макромолекул, включающий практически все структуры макромолекул (белки, ДНК, РНК и т. д.). Каждой структуре присвоен идентификационный номер (четырехбуквенный шифр, называемый PDB ID). Эти номера указаны в данной книге в подрисуночных подписях к каждому изображению структуры, взятой из базы PDB, что позволяет студентам и преподавателям исследовать приведенные структуры самостоятельно. В PDB описаны пространственные координаты каждого атома, положение которого было определено (многие приведенные в каталоге структуры еще не полностью описаны). В дополнительных файлах содержится информация о том, как была определена структура, и ее точность. По координатам атомов можно воспроизвести изображение макромолекулы с помощью специальной программы визуализации. Приветствуется посещение PDB студентами и исследование структур с помощью программы визуализации, на которую есть ссылка в базе. Кроме того, файлы со структурами макромолекул можно скачать и исследовать у себя на компьютере с помощью свободно распространяемых программ, таких как RasMol, Protein Explorer или FirstGlance HaJmol, они доступны по адресу www.umass.edu/microbio/rasmol.
Исследование структуры миоглобина позволило подобрать первые ключи к разгадке глобулярной структуры белка
Архитектура белка — третичная структура небольших глобулярных белков, II. Миоглобин.Первым прорывом в понимании трехмерной структуры белка стало исследование молекулы миоглобина методом рентгеноструктурного анализа, осуществленное Джоном Кендрю с коллегами в 1950-х гг. Миоглобин — это относительно небольшой белок мышечных клеток (Mr = 16 700), связывающий кислород. Его функция заключается как в хранении кислорода, так и в облегчении его диффузии в быстро сокращающихся мышечных тканях. Миоглобин состоит из одной полипептидной цепи, содержащей 153 аминокислотных остатка, последовательность которых известна, и гема (комплекса железа(II) с протопорфирином). Такой же гем есть в гемоглобине — белке эритроцитов, отвечающем за перенос кислорода. Именно с наличием гема связана темная красно-коричневая окраска миоглобина и гемоглобина. Особенно много миоглобина у морских млекопитающих — китов, тюленей и дельфинов, так что их мышцы имеют коричневый цвет. Механизм хранения и распределения миоглобина в мышцах этих животных позволяет им длительное время находиться под водой. Функции миоглобина и других глобинов более подробно рассматриваются в гл. 5.
На рис. 4-15 представлено несколько способов изображения пространственной структуры миоглобина, т. е. его третичной структуры. Красная группа внутри белка — это гем. Основная цепь молекулы миоглобина состоит из восьми относительно прямых участков α-спирали, прерываемых петлями, некоторые из них имеют структуру β-новоротов. Самая длинная α-спираль содержит 23 аминокислотных остатка, самая короткая — только 7; все спирали правые. Более 70% аминокислотных остатков миоглобина входят в состав α-спиральных участков. Рентгеноструктурный анализ позволил определить точное расположение всех R-групп, которые занимают практически все свободное место внутри свернутой молекулы белка.
Рис. 4-15. Третичная структура миоглобина кашалота (PDB ID 1МВO). Расположение молекулы одинаково на всех рисунках; гем выделен красным цветом. Смысл данной иллюстрации состоит не только в том, чтобы познакомить читателя с самой структурой миоглобина, но и продемонстрировать различные способы изображения белковых молекул, а) Полипептидный остов показан в виде ленточной модели (такой тип изображения был введен Джейн Ричардсон); хорошо видны участки со вторичной структурой, в частности α-спирали. б) На поверхности целой молекулы можно увидеть карманы в структуре глобулы, в которых могут связываться другие молекулы. в) Ленточная модель, на которой показаны боковые группы (выделены синим цветом) гидрофобных аминокислотных остатков (Leu, Ile, Val и Phe). г) Пространственная модель с изображением боковых групп; размер каждой сферы соответствует ван-дер-ваальсову радиусу атома. Гидрофобные остатки выделены синим цветом; большинство из них скрыто в глубине молекулы белка.
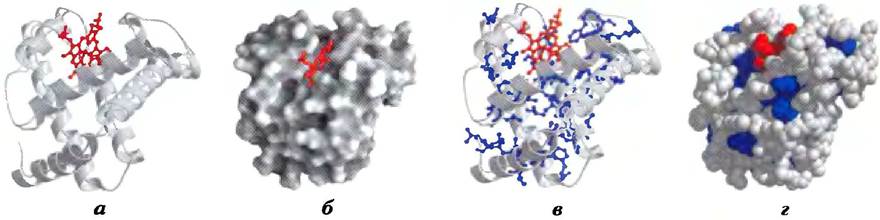
На основании анализа структуры миоглобина были сделаны многие важные выводы. Положение боковых цепей аминокислотных остатков связано с гидрофобными взаимодействиями, а значит, со стабильностью структуры белка. Большинство гидрофобных R-групп сосредоточены внутри молекулы миоглобина и поэтому не контактируют с водной средой. Все полярные R-группы миоглобина, за исключением двух, находятся на внешней поверхности молекулы в гидратированном состоянии. Молекула белка настолько компактна, что внутри глобулы хватает места лишь для четырех молекул воды. Наличие такого плотного гидрофобного ядра типично для глобулярных белков. В органической жидкости доля пространства, занимаемая атомами, составляет от 0,4 до 0,6, в типичном кристалле — от 0,70 до 0,78, т. с. близко к теоретическому максимуму, а в глобулярном белке — 0,75, что сравнимо со значением в кристалле. В таком тесном контакте между атомами слабые взаимодействия значительно усиливаются. Например, неполярные боковые цени аминокислотных остатков в гидрофобном ядре миоглобина расположены настолько близко, что действующие на коротких расстояниях ван-дер-ваальсовы взаимодействия вносят весьма заметный вклад в стабилизацию структуры.
Определение структуры миоглобина подтвердило некоторые предположения и выявило некоторые новые элементы вторичной структуры. Как предсказывали Полинг и Кори, все пептидные связи находятся в плоской трансконфигурации. Именно анализ структуры миоглобина впервые экспериментально подтвердил существование α-спиралей. Три из четырех остатков Pro в миоглобине расположены в изгибах последовательности. Четвертый остаток пролина находится внутри одной α-спирали, где образует петлю, необходимую для более плотной упаковки спиралей.
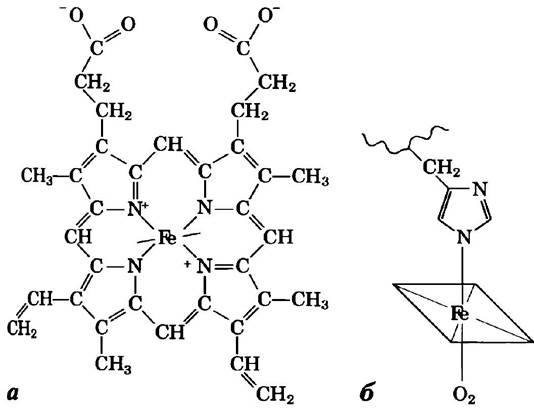
Гем имеет плоскую форму и находится в щели (кармане) на молекуле миоглобина. В центре гема расположен атом железа, две координационные связи которого направлены перпендикулярно к плоскости гема (рис. 4-16). Одна из этих связей соединена с R-группой остатка His в позиции 93, другая служит для связывания молекулы кислорода. Внутри кармана доступность гема для растворителя крайне ограничена. Это очень важно, поскольку свободный гем очень быстро окисляется, в результате чего ион Fe2+ превращается в Fe3+, а в такой форме гем не способен связывать кислород.
Рис. 4-16. Строение гема. Гемовая группа есть в миоглобине, гемоглобине, цитохромах и многих других белках. а) Гем представляет собой сложную полициклическую структуру (протопорфирин), с которой связан ион Fe2+. Атом железа имеет шесть координационных связей, четыре из которых расположены в плоскости молекулы порфирина и связывают атом Fe с ней; две другие связи направлены перпендикулярно порфириновому кольцу. б) В миоглобине и гемоглобине одна из связей, перпендикулярных порфириновому кольцу, занята атомом азота из остатка His. Другая связь свободна и служит для связывания молекулы кислорода.
Поскольку изучено много различных типов структуры миоглобина, у исследователей появилась возможность наблюдать структурные изменения, сопровождающие связывание кислорода или других молекул, что позволило впервые понять и продемонстрировать связь между структурой белка и его функцией. С тех пор аналогичные исследования были проведены для сотни белков. Сегодня результаты рентгеновской дифракции дополняются данными таких методов, как ЯМР-спектроскопия, что позволяет получить еще больший объем информации (доп. 4-5). В результате секвенирования геномов многих организмов (гл. 9) удалось идентифицировать тысячи генов, кодирующих белки с известной последовательностью, но с неизвестными пока функциями; это направление исследований стремительно развивается.
Дополнение 4-5. МЕТОДЫ. Методы определения трехмерной структуры белка
Метод рентгеноструктурного анализа
Положение атомов в кристаллической решетке можно определить, измеряя положение и интенсивность пятен (точек), высвечиваемых на фотопленке рентгеновскими лучами заданной длины волны после их дифракции на электронах этих атомов. Например, рентгеноструктурный анализ кристаллов хлорида натрия показывает, что ионы Na+ и Сlвместе образуют простую кубическую решетку. Подобным же образом можно определять расстояния между разными атомами в сложных органических молекулах, даже таких крупных, как белки. Но процедура анализа кристаллической структуры этих сложных молекул гораздо более трудоемкая, чем в случае простых солей. Если повторяющимся элементом кристаллической ячейки является такая большая молекула, как белок, то составляющие его многочисленные атомы дают тысячи дифракционных точек, и здесь нужны уже компьютерные методы обработки результатов.
Процесс дифракции рентгеновских лучей можно представить на примере получения изображения в световом микроскопе. Свет от точечного источника фокусируют на изучаемом объекте. Волны света рассеиваются объектом, а затем с помощью линз собираются вновь, создавая его увеличенное изображение. Минимальный размер объекта, который можно исследовать с помощью светового микроскопа, определяется длиной волны видимого света (от 400 до 700 нм). Объекты, размер которых меньше половины длины волны падающего света, не видны в микроскоп. Для изучения таких мелких объектов, как молекулы белка, нужно использовать рентгеновские лучи с длиной волны от 0,7 до 1,5 А (0,07-0,15 нм). Однако не существует линз, способных собирать рентгеновские лучи для получения изображения; в данном случае приходится напрямую изучать дифракционную картину и восстанавливать изображение с помощью математических методов.
Объем информации, извлекаемой из данных рентгеноструктурного анализа, зависит от упорядоченности структуры образца. Уже самые первые исследования дифракции фибриллярных белков волос и шерсти, имеющих достаточно регулярную организацию, позволили получить некоторые важные структурные параметры. Однако пучки фибриллярных белков не образуют кристаллов: молекулы белка вытянуты параллельно друг другу, но они не ориентированы в одном направлении. Более детальную информацию о трехмерной структуре молекул белка можно получить только на кристаллических образцах. Кристаллизация белков — это в некотором смысле искусство эксперимента, а нс наука, и структуры многих белков до сих пор неизвестны по той простой причине, что пока не удается получить их в кристаллическом виде. Практики сравнивают выращивание монокристалла белка с попыткой связать несколько шаров для боулинга с помощью скотча.
Проведение самого рентгеноструктурного анализа осуществляется в несколько этапов (рис. 1). Кристалл помещают между источником рентгеновского излучения и детектором и получают регулярный набор точек, которые называются рефлексами и возникают как результат дифракции пучка рентгеновских лучей на атомах, причем каждый атом в молекуле образца вносит свой вклад в расположение каждой точки. С помощью математического метода, основанного на преобразовании Фурье, из дифракционной картины получают карту электронной плотности белка. В данном случае компьютер выступает в роли «вычислительной линзы». Затем на основании карты электронной плотности создают модель структуры белка.
Рис. 1. Этапы работы при определении структуры миоглобина кашалота с помощью рентгеноструктурного анализа, а) Получение дифрактограммы кристаллического образца белка. б) Расчет карты электронной плотности белка на основании дифракционной картины. Здесь показана карта электронной плотности только части молекулы — ее гема, в) Участки наибольшей электронной плотности указывают на расположение атомных ядер; на основании этой информации воссоздается полная картина; модель структуры гема по карте его электронной плотности, г) Полная структура гемсодержащего миоглобина кашалота (РDB ID 2МВW).
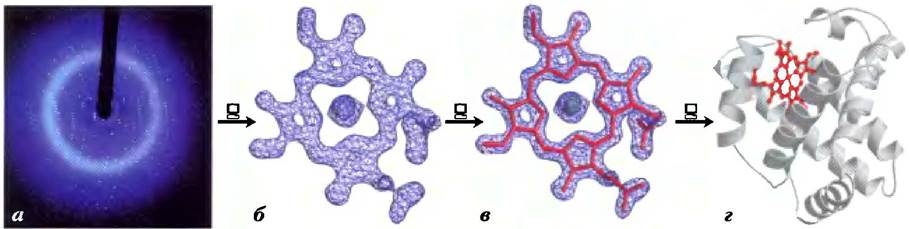
Джон Кендрю обнаружил, что рентгеновская дифрактограмма для кристаллического миоглобина (выделенного из мышц кашалота) носит сложный характер и состоит из 25 000 рефлексов. Компьютерный анализ этой картины осуществляли поэтапно. На каждом этапе разрешение улучшалось, и наконец, к 1959 г. было определено положение практически всех атомов белка (кроме атомов водорода). Аминокислотная последовательность белка, полученная методами химического анализа, совпадала с молекулярной моделью. С тех пор были определены структуры тысяч белков, некоторые из которых имеют гораздо более сложное строение, чем миоглобин.
Окружение атомов и молекул в кристалле, безусловно, отличается от окружения в растворе или в живой клетке. В модели, полученной при анализе кристалла, пространственные и временные параметры структуры усредняются. Рентгеноструктурный анализ почти ничего не говорит о конформациях, которые молекула белка может принимать в растворе. На конформацию белка в кристаллическом состоянии, в принципе, могут также оказывать влияние нефизиологические факторы, такие как случайные белок-белковые взаимодействия. Поэтому данные структурного анализа кристаллов сравнивают с данными, полученными другими методами (например, методом ЯМР, описанным ниже). При этом почти всегда выясняется, что структура молекулы, определенная с помощью рентгеноструктурного анализа, соответствует функциональной конформации белка. Рентгеноструктурный анализ может успешно применяться для изучения белков, размеры которых слишком велики для исследования методом ЯМР.
Метод ядерного магнитного резонанса (ЯМР)
Преимущество метода ЯМР состоит в том, что исследование осуществляется в растворе, тогда как рентгеноструктурный анализ применим только к тем объектам, которые удается кристаллизовать. Кроме того, методом ЯМР можно выявить динамические изменения в структуре белка, в том числе конформационные изменения, фолдинг и взаимодействие с другими молекулами.
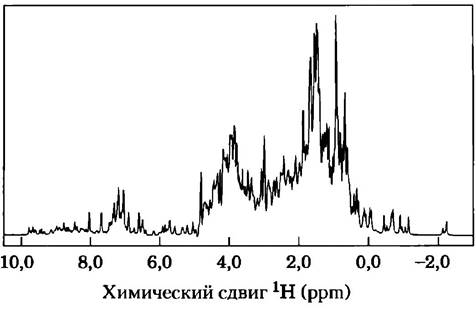
ЯМР напрямую связан с ориентацией ядерного магнитного момента — квантовомеханической характеристики атомных ядер. Только некоторые атомы, в том числе 1Н, 13С, 1N, ,9F и 31Р, имеют ядра, дающие сигнал ЯМР. (Все эти атомы имеют спиновое квантовое число 1/2- — Прим. ред.) Ядерный спин создает магнитный диполь. Если раствор, содержащий макромолекулы одного типа, поместить в сильное постоянное магнитное поле, то диполи выстраиваются в этом поле в одной из двух возможных ориентаций по направлению (с низким уровнем энергии) или против направления (с высоким уровнем энергии) поля. Диполи, выстроенные в магнитном поле, облучают под прямым углом короткими (-10 мкс) электромагнитными импульсами подходящей частоты (резонансной частоты, находящейся в диапазоне радиочастот). Ядра поглощают некоторое количество энергии и переходят в состояние с более высоким уровнем энергии, причем спектр поглощения содержит информацию о типах ядер и окружении, в котором они находятся. Данные многих подобных экспериментов усредняют, добиваясь максимального увеличения отношения сигнал/шум, и получают спектр ЯМР такого типа, как показан на рис. 2.
Рис. 2. Одномерный спектр ЯМР глобина морского червя. Этот белок очень близок по структуре к миоглобину кашалота; оба белка относятся к одному структурному семейству и отвечают за транспорт кислорода.
Особенно важную роль в методе ЯМР играют сигналы от атомов 1Н, что связано с их высокой чувствительностью и высоким содержанием в образцах. Однако в случае макромолекул протонный (1Н) спектр ЯМР может оказаться слишком сложным. Даже небольшой белок содержит сотни атомов 1Н, что обычно делает невозможным анализ одномерного спектра ЯМР. Структурный анализ белков стал возможен с развитием метода двумерной спектроскопии ЯМР (рис. 3). Этот метод позволяет определять зависимые от расстояния диполь- дипольные взаимодействия между спинами сближенных в пространстве атомов (измерение ядерного эффекта Оверхаузера (ЯЭО) в двумерном эксперименте NОЕSY), а также спин-спиновые взаимодействия атомов, связанных ковалентными связями (метод ТОСSY).
Рис. 3. Метод двумерной спектроскопии ЯМР для получения трехмерной структуры глобина (того же белка, что использовался для получения спектра на рис. 2).
Диагональ двумерного спектра ЯМР эквивалентна одномерному спектру. Пики, лежащие вне диагонали, соответствуют сигналам ЯЭО, возникшим в результате взаимодействия расположенных близко друг от друга атомов 1Н, сигналы которых в одномерном спектре могут быть достаточно удалены друг от друга. На рисунке (а) идентифицированы два таких взаимодействия; они показаны на рисунке (б) синими линиями (PDB ID 1VRF). Взаимодействие между метильной группой белка и водородом гема изображено с помощью трех линий. Метильная группа быстро вращается, так что все три ее атома водорода вносят одинаковый вклад во взаимодействие и в сигнал ЯМР. Эта информация использована для построения полной трехмерной структуры белка, приведенной на рисунке (в) (PDB ID IVRE). Множественные линии, изображающие полипептидный остов белка, демонстрируют серию структур, согласующихся с ограничениями межатомных расстояний, определенными методом ЯМР. Структурное сходство этого белка с миоглобином (рис. 1) очевидно. На обоих рисунках белки ориентированы одинаковым образом.
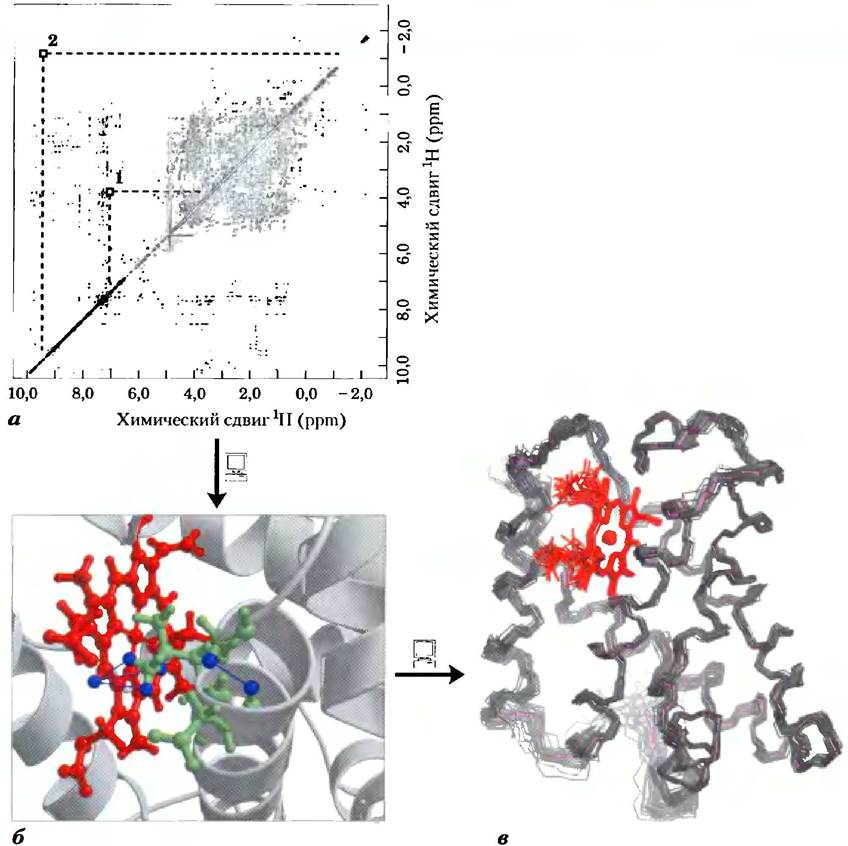
Построение полной трехмерной структуры на основании двумерного спектра ЯМР может оказаться очень трудоемкой задачей. Величины ЯЭО дают некоторую информацию о расстояниях между отдельными атомами, но, чтобы использовать эту информацию, каждый сигнал необходимо отнести к определенному атому. Какие сигналы ЯЭО соответствуют атомам, связанным ковалентными связями, можно узнать с помощью дополнительных экспериментов ТОСSY Определенные сигналы в ЯЭО-спектре удастся связать с наличием вторичной структуры белка, такой как α-спираль. Современные методы генной инженерии (гл. 9) позволяют создавать белки, содержащие изотопы 13С или 15N. Сигналы ЯМР, связанные с наличием этих атомов, в сочетании с сигналами, возникающими в результате их взаимодействия с 1Н, позволяют выявить сигналы отдельных протонов. Это легче сделать, если известна аминокислотная последовательность белка.
Для воспроизведения трехмерной структуры белка в компьютерную программу закладывают значения межатомных расстояний, а также такие известные геометрические параметры, как хиральность, ван-дер- ваальсовы радиусы, длины и углы связей. На основании этих данных компьютер создает серию структур, отражающих возможный диапазон конформационных состояний молекулы с учетом ограничений межатомных расстояний, полученных из данных ЯЭО (рис. 3. в). Некоторая неоднозначность структур, созданных на основе результатов ЯМР, частично отражает колебания внутри белковой молекулы в растворе, которые будут подробнее обсуждаться в гл. 5. Известная неопределенность проистекает также из ошибок эксперимента.
Обычно структуры белка, определенные с помощью рентгеноструктурного анализа и ЯМР, подтверждают друг друга. Иногда возможны расхождения в локализации боковых цепей отдельных аминокислотных остатков на поверхности белка, что часто связано с характером упаковки соседних молекул белка в кристалле. Эти два метода являются основным источником быстро растущего объема информации о структуре макромолекул в живых клетках.
Глобулярные белки имеют разные типы третичной структуры
Из наших знаний о третичной структуре сотен глобулярных белков ясно, что миоглобин является иллюстрацией всего лишь одного из множества возможных способов свертывания полипептидной цепи. В табл. 4-3 приведено относительное содержание аминокислотных остатков, входящих в состав α-спиральных участков и участков с β-конформацией, для нескольких небольших глобулярных белков, состоящих из одной полипептидной цепи. Каждый из этих белков имеет характерную структуру, адаптированную для выполнения специфической биологической функции, однако у них есть важные общие свойства. Каждый белок образует компактную структуру, причем гидрофобные боковые цепи ориентированы внутрь глобулы (подальше от воды), а гидрофильные боковые цепи расположены на поверхности. Все структуры стабилизированы с помощью множества водородных связей и некоторых ионных взаимодействий.
Таблица 4-3. Приблизительное содержание аминокислотных остатков в α-спиралях и участках с β конформацией в некоторых одноцепочечных белках
Белок (общее число остатков) |
Остатки α-Спирали |
(%)* β-Конформация |
Химитрипсин (247) |
14 |
45 |
Рибонуклеаза (124) |
26 |
35 |
Карбоксипентидаза (307) |
38 |
17 |
Цитохром с(104) |
39 |
0 |
Лизоцим (129) |
40 |
12 |
Миоглобин (153) |
78 |
0 |
* Участки полипептидных цепей, не образующих ни α-спирали, ни β-конформацию, представляют собой изгибы цепи и нерегулярно скрученные или вытянутые отрезки. Участки α-спиралей и β-конформации иногда имеют слегка искаженные размеры и геометрию, отличающиеся от соответствующих нормальных параметров.
Для первого знакомства с очень сложной третичной структурой глобулярных белков, размеры которых гораздо больше размеров миоглобина, полезно обратить внимание на несколько типов структур, встречающихся в самых разных, часто вовсе не связанных между собой белках. Трехмерную структуру типичного глобулярного белка можно представить, как набор полипептидных сегментов, уложенных в виде α-спиралей или β-слоев и соединенных друг с другом связующими последовательностями. Следующим шагом является описание того, как эти сегменты соединены друг с другом и как организованы связующие последовательности.
Понимание трехмерной структуры белка в целом строится на анализе структуры отдельных его участков. Мы начнем с определения двух важных терминов, использующихся для описания отдельных участков или элементов структуры полипептидной цепи, а затем обратимся к закономерностям укладки (фолдинга) белка.
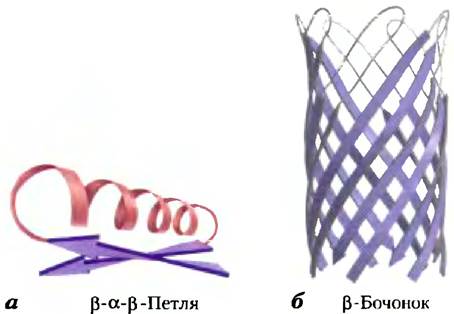
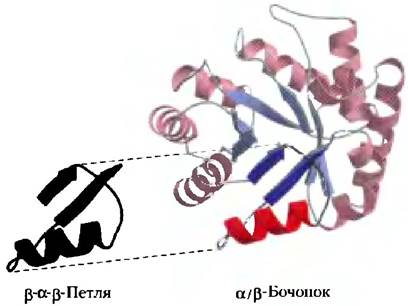
Первый термин — это супервторичная структура белка, иначе называемая белковым мотивом; эта структура определяет устойчивое взаиморасположение определенных элементов вторичной структуры и связующих их последовательностей. Приведенные термины синонимичны, хотя в литературе наблюдается некоторая путаница с их употреблением. Белковые мотивы могут быть очень простыми, например, два элемента вторичной структуры, расположенные друг напротив друга, и составлять лишь малую часть белка. Пример таких элементов — β-α-β-петли (рис. 4-17, а). Мотив может быть очень замысловатой структуры и содержать множество участков белка, упакованных в единую структуру, например, β-бочонок (рис. 4-17, б). Иногда целый белок может являться одним большим мотивом. Этот термин отражает все преимущества представления белка в виде отдельно упакованных участков и хорош для описания таких участков. Участок, определенный как мотив, иногда бывает устойчивой самостоятельной структурой. Мы уже сталкивались с одним хорошо изученным мотивом — суперспиралью α-кератина, которая встречается и в ряде других белков. Обратите внимание, что мотив — это не промежуточный уровень структуры белка между вторичной и третичной структурами. Это участок укладки, который описывает малую часть белка или отдельную полипептидную цепочку. Синоним «супервторичная структура» несколько запутывает, гак как подразумевает наличие определенной иерархии.
Рис. 4-17. Мотивы, а) Простой мотив — β-α-β-петля. б) Более сложный мотив — β-бочонок. Этот β-бочонок является единственным доменом α-гемолизина (токсина, уничтожающего бактерии путем создания пор в клеточных мембранах) из бактерии Staphylococcus aureus (PDB ID 7AHL).
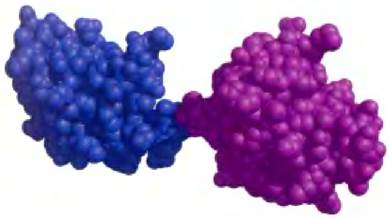
Второй термин, используемый для описания структуры белка, это домен. По определению Джейн Ричардсон, данному в 1981 г., домен — это часть полипептидной цепи, которая стабильна вне зависимости от остальной цепи или может перемещаться по отношению к целому белку как самостоятельная единица. Полипептиды, имеющие в своем составе несколько сотен аминокислотных остатков, часто состоят из двух или более доменов, которые могут различаться по функциям. Во многих случаях домены большого белка сохраняют свою нативную трехмерную структуру даже при отделении от другой части белка, например, в результате протеолитического расщепления. Иногда домены в мультидоменных белках удается различить как отдельные структурные единицы (рис. 4-18), но чаще этого сделать нельзя из-за тесного контакта между ними. Отдельные домены часто выполняют различные функции, такие как связывание небольших молекул или взаимодействие с другими белками. Маленькие белки обычно состоят из одного домена (т. е. домен представляет собой весь белок).
Рис. 4-18. Структурные домены в полипептидной цепи тропонина С (РDВ ID 4TNC). Данный кальций-связывающий белок мышечной ткани содержит отдельные домены для связывания кальция, выделенные на рисунке разными цветами.
Сворачивание белков (фолдинг) подчиняется ряду физических и химических закономерностей. Перечислим некоторые из них, поскольку они позволят определить несколько простых белковых мотивов.
1. Гидрофобные взаимодействия вносят значительный вклад в устойчивость структуры белка. Необходимость спрятать R-группы гидрофобных аминокислот, чтобы исключить их контакт с водой, влечет за собой возникновение, по крайней мере, двух уровней вторичной структуры. Такие два уровня могут создаваться простыми мотивами, например, β-α-β-петлями (рис. 4-17, а).
2. Если в белках существуют как α-спирали, так и β-слои, то они обычно расположены в разных частях структуры. Это правило объясняется тем, что полипептидный участок с β-конформацией (рис. 4-6) не может участвовать в образовании водородных связей с находящейся рядом α-спиралью.
3. Полипептидные участки, расположенные рядом в последовательности аминокислот, обычно оказываются по соседству и в свернутом белке. Хотя удаленные участки последовательности в результате образования третичной структуры также могут соприкасаться, это происходит нечасто.
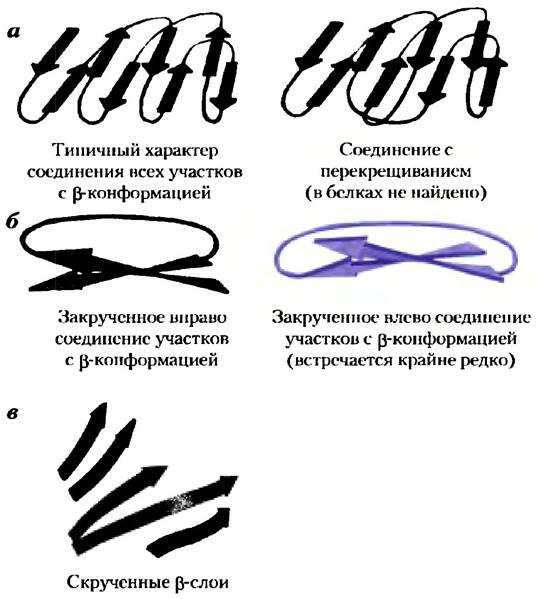
4. Связующие последовательности между элементами вторичной структуры не пересекаются и не образуют узлов (рис. 4-19, а).
5. β-Конформация оказывается более устойчивой, если ее отдельные участки слегка скручены вправо. Это влияет на расположение как обоих листов по отношению друг к другу, так и полипептидного связующего отрезка между ними. Например, два параллельных β-слоя должны перекрещиваться (рис. 4-19, б). В принципе, такая перекрестная структура может быть повернута как вправо, так и влево, но в белках почти всегда реализуется правая конформация. Правая связующая последовательность обычно короче левой и изогнута под меньшим углом, поэтому она легче образуется. Скрученность β-слоев, кроме того, приводит к характерным изгибам структуры, образующейся при контакте большого числа таких участков. Примерами могут служить β-бочонок (рис. 4-17, б) и скрученные β-слои (рис 4-19, в), которые образуют ядро многих более крупных структур.
Рис. 4-19. Устойчивые типы укладки полипептидной цепи в белках, а) Связь между участками цепи с β-конформацией в уложенных β-слоях. Цепи показаны с одного конца, изгибы структуры не отражены. Более толстыми линиями обозначены связующие последовательности, расположенные ближе к читателю, более тонкими линиями — те, что расположены на дальней стороне β-слоя. Связующие последовательности с каждой стороны β-слоя (как с ближней к читателю, так и с дальней от него стороны) не пересекаются между собой. 6) Участки с β-конформацией скручиваются, поэтому связывающие их последовательности обычно закручены вправо. Закрученные влево структуры должны образовывать более острые углы, поэтому они труднее образуются, в) Скрученные β-слои — это структура домена фотолиазы (фермента, ответственного за репарацию некоторых типов повреждений ДНК) из Е. coli (PDB ID 1DNP). Соединяющие петли не показаны для наглядности изображения укладки β-слоев.
Более сложные белковые мотивы могут строиться из более простых в соответствии с данными правилами. Например, серия β-α-β-петель, организованных так, что участки с β-конформацией формируют бочонок, образует очень устойчивый и распространенный мотив, называемый αβ- бочонком (рис. 4-20). В этой структуре каждый из параллельных β-слоев связан с соседним с помощью α-спирального участка. Все связующие последовательности закручены вправо. αβ-Бочонок обнаружен во многих ферментах, причем участок связывания кофактора или субстрата часто находится в кармане, расположенном у одного из концов бочонка. Обратите внимание, что элементы со сходным характером укладки полипептидной цепи считают одинаковыми мотивами, даже если входящие в их состав α-спирали и β-слои имеют разную протяженность.
Рис. 4-20. Построение более крупного мотива из более мелких. β-Бочонок - это распространенный мотив из повторяющихся мотивов β-α-β-петель. Изображен домен гликолитического фермента пируваткиназы кролика (РDВ ID 1РKN).
Белковые мотивы — основа классификации белковых структур
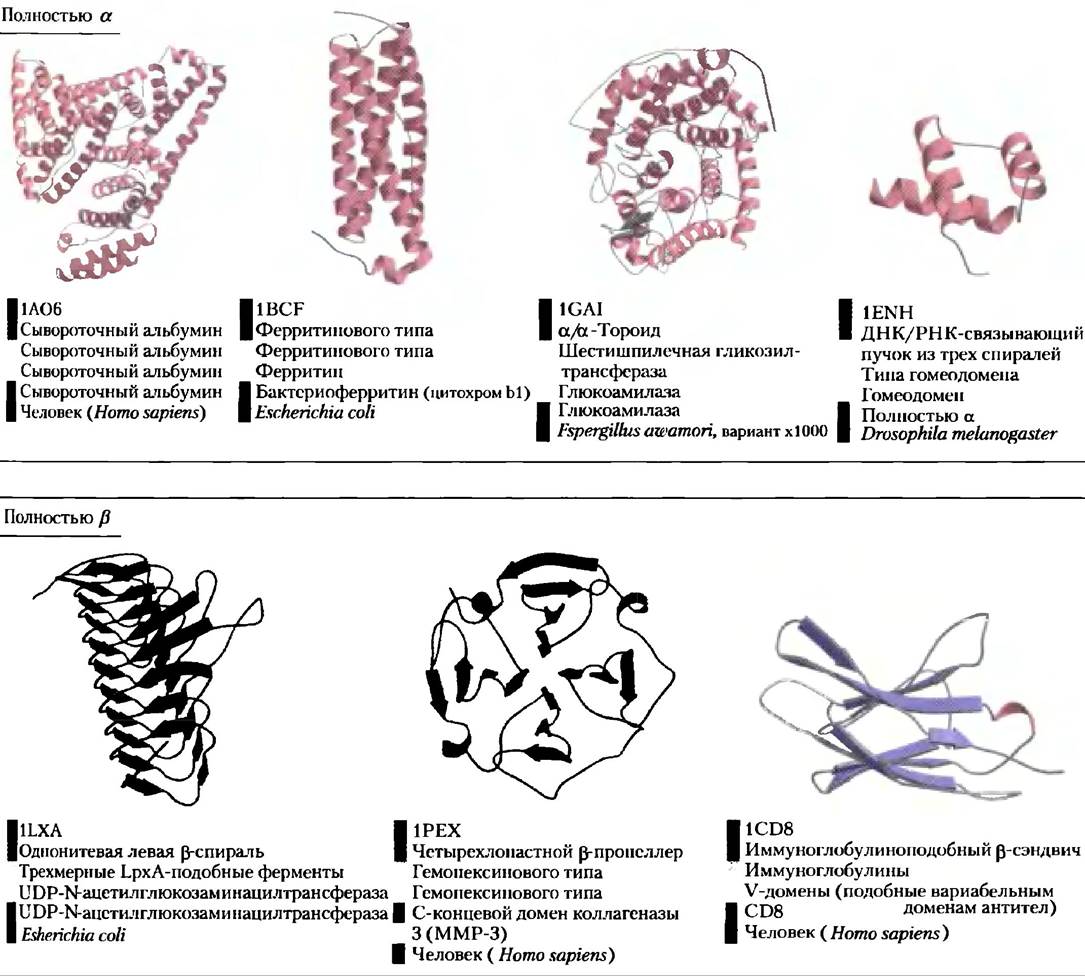
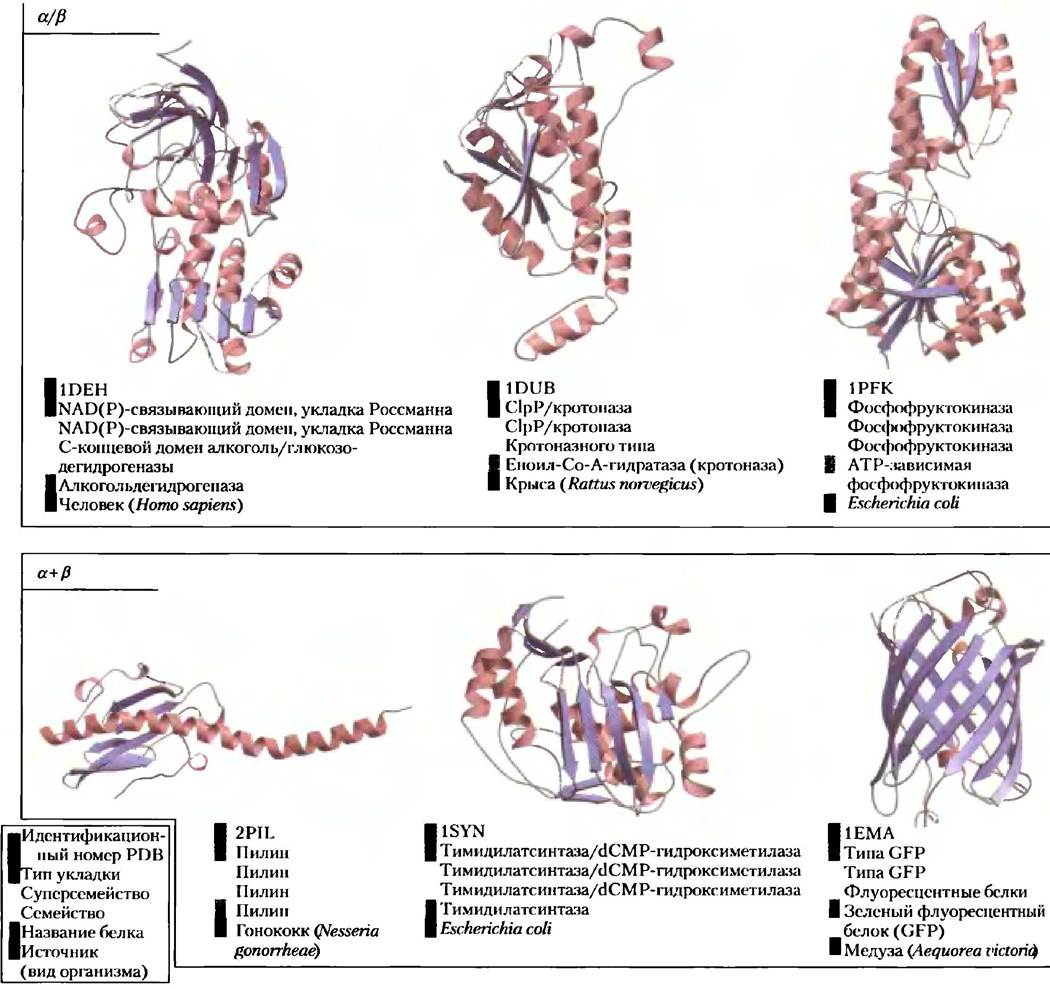
Архитектура белка — третичная структура больших глобулярных белков, IV. Структурная классификация белков. Как вы могли заметить, изучение третичной структуры белка несколько упрощается, если принять во внимание наличие во многих белках характерных элементов (подструктур). Развивая такой подход к анализу белков, ученые организовали содержимое баз данных о белковых структурах в соответствии с иерархическими уровнями организации белков. База данных SCOP (StructuralClassification of Proteins), содержащая информацию о структурной классификации белков, является примером этого крайне важного направления в биохимии. Для самого высокого уровня классификации база данных SCOP (http://scop.mrc-lmb.cam.ac.uk/scop/) использует уже широко известную схему, в соответствии с которой все белковые структуры делятся на четыре класса: полностью α, полностью β, α/β (участки с α- и β-структурой рассеян, по последовательности или чередуются) и α + β (участки с α- и β-структурой каким-то образом разделены и группируются с себе подобными). К каждому классу относятся от десятков до сотен различных типов укладки, имеющих различные типы подструктуры, и число их постоянно растет. Некоторые типы подструктуры встречаются очень часто, другие пока обнаружены в одном единственном белке. На рис. 4-21 приведены примеры белковых мотивов, встречающихся в белках четырех названных классов. И это лишь небольшая часть из сотен известных мотивов. Но количество вариантов укладки белковой цепи не бесконечно. Хотя скорость появления новых структур белков постоянно растет, рост числа новых идентифицируемых мотивов сокращается. Всего в белках может содержаться не долее 1000 различных типов укладки или мотивов, рис. 4-21 видно, как могут быть организован белки с теми или иными мотивами. Два верхних уровня организации белковых структур — классы и мотивы — касаются исключительно структуры белка. Ниже этих уровней классификация строится на эволюционных закономерностях.
Рис. 4-21. Классификация белков на основании структурных мотивов. Здесь представлена лишь небольшая часть из сотен известных устойчивых мотивов. Все они разделены на четыре класса: только α, только β, α/β и α + β. Кроме того, приводятся данные классификации SCOP (http://scop.mrc-lmb.cam.ac.uk/scop/) (см. цветовой код) и идентификационные номера из банка данных PDB (www.rcsb.org) (верхняя подпись у каждой структуры). Изображенный на рис. 4-20 αβ-бочонок является еще одним примером белка со структурой α/β.
Существует множество примеров доменов или мотивов, встречающихся в самых разных белках. Это означает, что третичная структура белка более консервативна, чем его первичная структура. Таким образом, сопоставление белковых структур может способствовать получению важной информации о ходе эволюции. Белки, имеющие значительное сходство первичной структуры и (или) очевидное сходство структуры и функций, относят к одному и тому же семейству белков. Внутри семейства белков обычно прослеживается явное эволюционное родство. Например, семейство глобинов содержит большое количество белков, каждый из которых похож на миоглобин как по структуре, так и по аминокислотной последовательности (см. доп. 4-5 и гл. 5). Белки двух или нескольких семейств, имеющиъ мало сходства в первичных структурах, но содержащие одинаковые пространственные структурные мотивы и близкие по функциям, объединяют в суперсемейства. Возможно, что между семействами внутри одного суперсемейства существует определенное эволюционное родство, даже если время и функциональные различия (а значит, различное адаптивное воздействие) стерли сходство последовательностей. Если белки одного семейства могут быть найдены во всех трех высших таксонах живых организмов — в бактериях, археях и эукариотах, это говорит об очень древнем происхождении семейства. Другие семейства могут распространяться лишь на небольшую группу организмов, что говорит о более позднем происхождении. Прослеживая историю структурныхмотивов с помощью классификационных баз данных, таких как SCOP, можно получить много полезной информации относительно эволюционных связей белков. База данных SCOP пополняется вручную и служит для размещения белков соответствии с их происхождением на основании консервативных элементов структуры.
Структурные мотивы очень важны для описания семейств и суперсемейств белков. Совершенствование системы классификации неизбежно привело к выявлению новых функциональных связей. Учитывая важнейшую роль белков для живых организмов, подобный сравнительный анализ структур может пролить спет на многие аспекты биохимии — от эволюции отдельного белка до эволюции целых метаболических путей.
Некоторые ресурсы и базы данных, работающие в режиме реального времени, дополняют SCOP. База данных CATH (class, architecture, topology, and homologous superfamily)позволяет поместить белок в банк белковых структур РDБ в соответствии с четырехуровневой иерархией. Другие программы позволяют пользователю ввести в программу интересующий его белок и найти в банке PDB другие белки, которые имеют с ним определенное сходство. Среди таких программ можно назвать VAST (vector alignment search tool), CE (combinatorial extension of the optimal paths) и FSSP (fold classification based on structure-structure alignment of proteins).
Четвертичная структура белка варьирует от простых димеров до больших комплексов
Архитектура белка — четвертичная структура.
Многие белки состоят из нескольких полипептидных субъединиц, что может служить для нескольких целей. Некоторые мультисубъединичные белки несут регуляторную функцию; связывание небольших молекул может влиять на взаимодействие между субъединицами и вызывать значительные изменения в активности белка в ответ на незначительные изменения концентрации субстратов или регуляторных молекул (гл. 6). В других случаях отдельные субъединицы могут иметь разные, но связанные между собой функции, например, катализ и регуляцию. Комплексы фибриллярных белков, обсуждавшиеся ранее в данной главе, или белки оболочки вируса выполняют структурную функцию. Некоторые очень крупные белковые комплексы участвуют в осуществлении многостадийных процессов. Примером являются рибосомы, осуществляющие биосинтез белка и состоящие из десятков белковых субъединиц и нескольких молекул РНК.
Мультисубъединичные белки часто называют мультимерами. Мультимер, состоящий всего из нескольких субъединиц, называют олигомером. Если мультимерный белок содержит различные субъединицы, то его общая структура может быть асимметричной и достаточно сложной. Однако большинство белков состоят из идентичных субъединиц или повторяющихся групп неидентичных субъединиц и обычно имеют симметричное строение. Как говорилось в гл. 3, повторяющиеся структурные единицы в таких мультисубъединичных белках называют протомерами, вне зависимости от того, идет ли речь о единственной субъединице или о группе субъединиц. Иногда для обозначения отдельных субъединиц протомера используют греческие буквы.
Первым олигомерным белком, чью трехмерную структуру удалось установить, был гемоглобин (64 500), состоящий из четырех полипептидных цепей и четырех гемов с атомами железа в форме Fe2+ (рис. 4-16). Белковая часть гемоглобина, называемая глобином, состоит из двух α-субъединиц (по 141 остатку в каждой) и двух β-субъединиц (но 146 остатков). Обратите внимание, что в данном случае обозначения а и β не имеют отношения к вторичной структуре. Поскольку гемоглобин в четыре раза крупнее миоглобина, на определение его трехмерной структуры с помощью рентгеноструктурного анализа было потрачено гораздо больше времени и сил. Структура гемоглобина была расшифрована Максом Перутцем, Джоном Кендрю и их коллегами в 1959 г. Субъединицы гемоглобина расположены симметричными парами (рис. 4-22): в каждой паре есть одна а- и одна β-субъединица. Таким образом, гемоглобин можно считать либо тетрамером, либо димером αβ-протомеров.
Макс Перутц, 1914-2002 (слева), Джон Кендрю, 1917-1997

Идентичные субъединицы мультимерных белков обычно организованы в виде некоторого числа симметричных структур. Для обсуждения этих структур необходимо объяснить некоторые термины, используемые при описании симметрии. Олигомеры могут иметь поворотную симметрию или винтовую (спиральную) симметрию. Это означает, что одну субъединицу можно наложить на другую (до точного совпадения) либо путем вращения вокруг одной или нескольких осей, либо в результате поворота и сдвига (поворота по спирали). В белках с поворотной симметрией субъединицы располагаются вокруг оси вращения и образуют замкнутые структуры. Белки со спиральной симметрией образуют более открытые структуры, в которых субъединицы выстраиваются одна за другой по спирали.
Существует несколько типов поворотной симметрии. Простейшим вариантом является симметрия, описываемая циклическими группами Сn; при этом симметричные объекты накладываются в результате поворота вокруг одной оси (рис. 4-23, а). (С означает циклическая, а п указывает на число субъединиц, связанных осью вращения.) Сама ось называется осью вращения n-го порядка. Таким образом, αβ-протомеры гемоглобина (рис. 4-22) связаны с осью симметрии С2. Несколько более сложной является симметрия, описываемая группами Dn (их иногда называют диэдрическими). В этом случае оси второго порядка пересекают ось n-го порядка под прямыми углами (рис. 4-23, б). Белок с симметрией Dn имеет 2n протомеров.
Рис. 4-22. Четвертичная структура дезоксигемоглобина (РDВ ID 2ННВ). Рентгеноструктурный анализ дезоксигемоглобина (гемоглобин без связанных с гемом молекул кислорода) позволяет понять, как расположены в пространстве четыре субъединицы белка, а) Ленточная модель. б) Модель поверхности. α-Субъединицы показаны серым цветом, β-субъединицы — оттенками синего. Обратите внимание, что гемовые группы (красного цвета) удалены друг от друга на относительно большое расстояние.
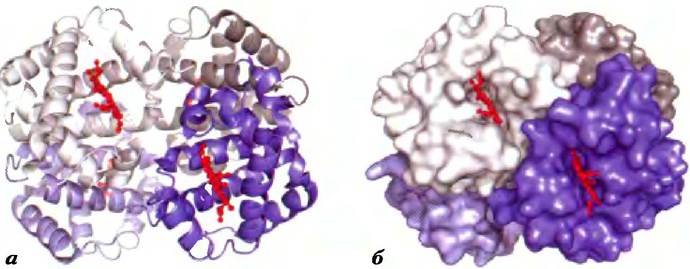
Рис. 4-23. Вращательная симметрия в белках, а) В случае поворотной симметрии типа С субъединицы мо но совместить путем поворота вокруг одной оси n-порядка, где n — это число связанных таким образ субъединиц. Оси изображены в виде черных лик» цифры указывают порядок оси. Показаны только две возможных структур с симметрией Сn. б) В случае поворотной симметрии типа D субъединицы могут быть совмещены в результате вращения относительно одной или двух осей, причем одна из двух — это ось 2-го порядка. Наиболее часто встречается симметрия типа в) Икосаэдрическая симметрия. Для совмещения всех треугольных граней икосаэдра необходимо вращения вокруг одной или большего числа осей симметрии - 3 и 5-го порядка. Слева показан вид икосаэдра перпендикулярно оси каждого типа.
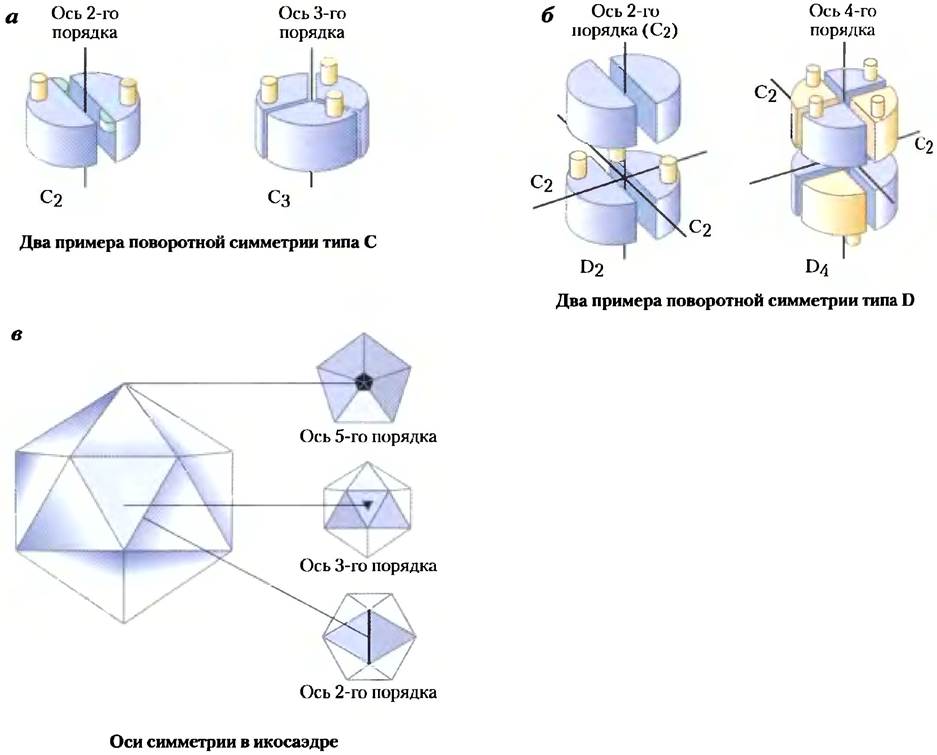
Белки с симметрией Сn или Dn встречаются наиболее часто. Возможны и другие типы поворотной симметрии, но лишь немногие из них обнаружены в белках. В качестве примера можно привести икосаэдрическую симметрию. Икосаэдр — это правильный двенадцативершинник, имеющий 20 равносторонних треугольных граней (рис. 4-23, в). Каждую грань можно совместить с остальными путем поворота вокруг одной или большего числа осей трех видов. Такую структуру очень часто имеет оболочка вируса - капсид. Возбудитель полиомиелита человека (полиовирус) имеет капсид такого строения (рис. 4-24, а). Каждая треугольная грань составлена из трех протомеров, содержащих по одной копии четырех различных полипептидных цепей, три из которых находятся на внешней поверхности капсида. Шестьдесят протомеров образуют 20 сторон оболочки в форме икосаэдра, внутри которой хранится генетический материал (РНК).
Рис. 4-24. Капсиды вирусов, а) Полиовирус (PDB ID 2PLV), как он представлен в базе структурной вирусологии VIPER. Белки на поверхности полиовируса собраны в виде икосаэдра с диаметром 300 А. Икосаэдрическая симметрия — один из видов вращательной симметрии (рис. 4-23, в). Слева представлено изображение поверхности капсида полиовируса. Изображение справа дано с малым разрешением; субъединицы белков капсида раскрашены, чтобы показать икосаэдрическую симметрию. 6) Вирус табачной мозаики (PDB ID 1VTM). Этот вирус имеет форму палочки (как показано на электронной микрофотографии) длиной 3000 А и диаметром 180 А и обладает спиральной симметрией.
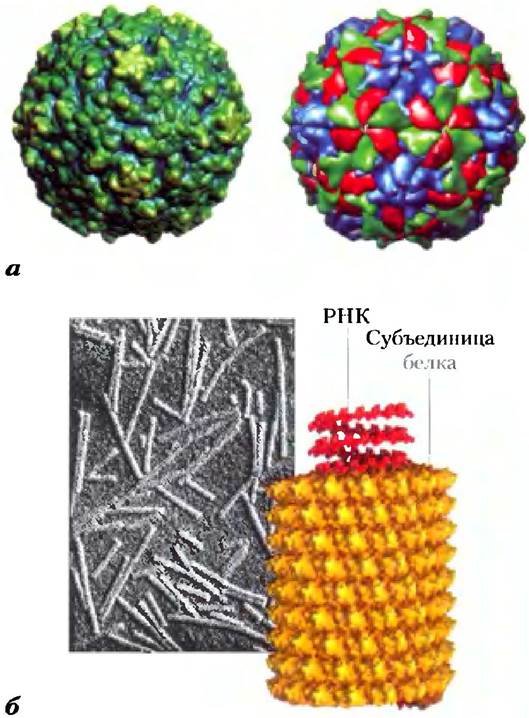
Винтовая симметрия, встречающаяся в олигомерных белках, тоже была обнаружена в капсидах. Вирус табачной мозаики представляет собой правозакрученную спираль, состоящую из 2130 идентичных субъединиц (рис. 4-24, б). Внутри такой цилиндрической структуры содержится РНК вируса. Белки, состоящие из расположенных по спирали субъединиц, также могут формировать протяженные нитевидные структуры, такие как актиновые филаменты в мышцах (рис. 5-28).
Краткое содержание раздела 4.3 Третичная и четвертичная структуры белка
■ Третичная структура представляет собой полную пространственную структуру полипептидной цепи. На основании третичной структуры белки делятся на два больших класса — фибриллярные и глобулярные.
■ Фибриллярные белки, обычно выполняющие структурные функции, как правило, включают простые повторяющиеся элементы вторичной структуры.
■ Глобулярные белки имеют гораздо более сложную третичную структуру и часто в одной и той же полипептидной цепи содержат элементы разных типов вторичной структуры. Миоглобин — это первый глобулярный белок, третичная структура которого была определена методом рентгеноструктурного анализа.
■ Сложное строение глобулярных белков можно изучать, анализируя устойчивые подструктуры — супервторичные структуры, или мотивы. Для построения тысяч известных белковых последовательностей используется лишь несколько сотен мотивов. Участки полипептидной цепи, способные образовывать устойчивые и независимые структуры, называют доменами.
■ Четвертичная структура белка возникает в результате взаимодействия субъединиц мультисубъединичного (мультимерного) белка или крупных белковых комплексов. Некоторые мультимерные белки построены из повторяющихся единиц, состоящих из одной субъединицы или группы субъединиц (протомеров). Протомеры обычно связаны между собой поворотной или винтовой симметрией.