ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 1. ОСНОВЫ БИОХИМИИ СТРОЕНИЕ И КАТАЛИЗ - 2011
ЧАСТЬ I. СТРОЕНИЕ И КАТАЛИЗ
9. ТЕХНОЛОГИЯ НА ОСНОВЕ ИНФОРМАЦИИ ИЗ ДНК
Вопросы и задачи
1. Клонирование.
При соединении двух или более фрагментов ДНК ученые могут подбирать последовательность в месте слияния множеством хитроумных способов, рассматриваемых в следующих упражнениях.
а) Изобразите структуру каждого конца линейного фрагмента ДНК, образуемого при рестрикции ферментом EсоRI (включая последовательности, остающиеся от сайта узнавания EсоRI).
б) Изобразите структуру, получающуюся в результате реакции этой концевой последовательности с ДНК-полимеразой I и четырьмя дезоксинуклеозидтрифосфатами (см. рис. 8-33).
в) Приведите последовательность, получающуюся в месте стыковки двух концов с полученной в (б) структурой при их сшивке (см. рис. 25-17).
г) Изобразите структуру, получающуюся, если на структуру из (а) воздействовать нуклеазой, разрушающей только одноцепочечную ДНК.
д) Приведите последовательность стыка при сшивке конца со структурой (б) с концом со структурой (г).
е) Изобразите структуру конца линейного фрагмента ДНК, образованного при рестрикции ферментом РvuII (рассмотрите последовательности, остающиеся от последовательности распознавания Pvull).
ж) Приведите последовательность стыка при сшивке конца со структурой (б) с концом со структурой (е).
з) «Синтезируйте» короткий фрагмент двуцепочечной ДНК с любой последовательностью на ваше усмотрение. На основе этого синтетического фрагмента и методов из пунктов от (а) до (ж) разработайте схему удаления из молекулы ДНК сайта рестрикции EcoRI и встраивания нового сайта рестрикции ВаmHI примерно в том же месте (см. рис. 9-3).
и) Предложите четыре различных коротких синтетических фрагмента двухнитевой ДНК, которые позволили бы сшить структуру (а) с фрагментом ДНК, образованным при рестрикции ферментом Pstl. В одном из этих фрагментов сконструируйте последовательность, так чтобы в конечном соединении были последовательности распознавания как EcoRI, так и Pst\. Во втором и третьем фрагментах сконструируйте последовательности так, чтобы соединение содержало последовательность распознавания либо EcoRI, либо PstI соответственно. В четвертом фрагменте разработайте последовательность так, чтобы в конечном соединении не было ни сайта рестрикции EcoRI, ни Pstl.
2. Отбор рекомбинантных плазмид.
При клонировании фрагмента чужеродной ДНК в плазмиде часто бывает полезно вставлять фрагмент на участке, нарушающем целостность селектируемого маркера (например, ген устойчивости к тетрациклину плазмиды pBR322). Потерю своей функции нарушенным геном можно использовать для обнаружения клонов, содержащих рекомбинантные плазмиды с чужеродной ДНК В случае вектора на основе бактериофага λ в этом нет необходимости, поскольку можно легко отличить векторы с огромными фрагментами чужеродной ДНК от тех, у которых их нет. Как такие рекомбинантные векторы идентифицируют?
3. Клонирование ДНК.
Плазмидный клонирующий вектор pBR322 (см. рис. 9-4) разрезается эндонуклеазой рестрикции Pstl. Взятый из эукариотического генома фрагмент ДНК (также полученный разрезанием Pstl) вставляется в приготовленный вектор и сшивается. Раствор сшитых таким образом ДНК затем используется для трансформации бактерий, а бактерии с плазмидами отбираются выращиванием в присутствии тетрациклина.
а) Кроме искомой рекомбинантной плазмиды какие еще типы плазмид можно обнаружить среди трансформированных бактерий, устойчивых к тетрациклину? Как их можно различить?
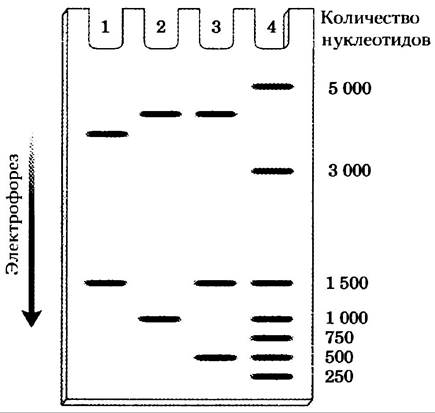
б) Фрагмент клонируемой ДНК имеет длину в 1000 п. н., а сайт EcoRI расположен на расстоянии в 250 п. н. от одного из концов. Три разных рекомбинантных плазмиды разрезают EсоRI и анализируют гель-электрофорезом, дающим приведенные паттерны. Что можно сказать о клонируемой ДНК на основании каждого из паттернов? Имейте в виду, что в рВR322 участки рестрикции PStI и EсоRI находятся на расстоянии 750 п. н. друг от друга. Вся плазмида целиком без клонируемой вставки имеет длину 4361 п. н. По маркерам размеров на дорожке 4 можно оценить длину нуклеотидного фрагмента.
4. Идентификация гена белка с известной аминокислотной последовательностью.
Используя рис. 27-7 для трансляции генетического кода, предложите ДНК-зонд, который позволил бы идентифицировать ген белка со следующей N-концевой аминокислотной последовательностью. Зонд должен иметь длину от 18 до 20 аминокислот — размер, который обеспечивает надлежащую специфичность, если имеется достаточная гомология между геном и зондом.
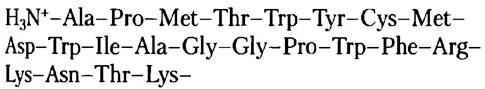
5. Создание диагностических тестов генетических заболеваний.
Болезнь Хантингтона (БХ) является наследственным нейродегенеративным нарушением, характеризующимся постепенным, необратимым ухудшением психических, двигательных и умственных способностей. Симптомы обычно появляются в среднем возрасте, тем не менее приступ болезни может начаться практически в любом возрасте. Болезнь может длиться от 15 до 20 лет. Сейчас молекулярная основа заболевания становится более понятной. Генетическая мутация, лежащая в основе БХ, была обнаружена в гене, кодирующем белок (Мr = 350 000) с неизвестной функцией. У людей, не страдающих БХ, участок гена, кодирующий N-конец белка, состоит из последовательностей кодонов САG (соответствует глутамину), повторяющихся от 6 до 39 раз подряд. У людей с синдромом БХ в зрелом возрасте этот кодон, как правило, повторяется от 40 до 55 раз, а у людей, страдающих БХ с детства, — более 70 раз. На основании длины этого простого трехнуклеотидного повтора можно сказать, разовьется ли у человека БХ, и примерно в каком возрасте появятся первые симптомы.
Внизу показана небольшая часть N-концевой кодирующей последовательности гена БХ, состоящего из 3143 кодонов. Нуклеотидная последовательность ДНК выделена черным цветом, аминокислотная последовательность, соответствующая гену, показана синим цветом, а САG-повтор затенен. Используя рис. 27-7 для трансляции генетического кода, опишите в общих чертах тест на БХ на основе ПЦР, который можно было бы осуществить в образце крови. Примите, что праймер для ПЦР должен иметь длину 25 нуклеотидов. По традиции, если не оговаривается иначе, последовательность ДНК, кодирующая белок, изображается кодирующей цепочкой (последовательность, идентичная мРНК, которая транскрибируется из гена) сверху, так чтобы она читалась от 5'-конца к 3'-концу слева направо.
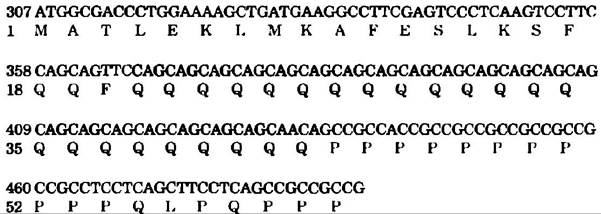
6. Использование ПЦР для обнаружения кольцевых молекул ДНК.
У реснитчатых одноклеточных организмов иногда удаляется сегмент геномной ДНК Удаление является генетически запрограммированной реакцией, связанной со скрещиванием клеток. Ученые предполагают, что ДНК удаляется в процессе сайт-специфичной рекомбинации, ДНК на обоих концах сегмента, которые скрепляются вместе, а удаленная ДНК появляется в виде кольцевой молекулы в качестве продукта реакции.
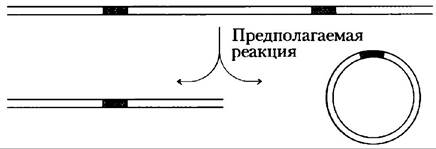
Подумайте, как можно использовать полимеразную цепную реакцию (ПЦР) для обнаружения циклической формы, удаленной ДНК в экстракте одноклеточных организмов.
7. Светящиеся растения.
Выращенные на обычной садовой почве в условиях нормального полива растения, несущие ген зеленого флуоресцентного белка (см. рис. 9-15, а), светятся в темноте, а растения, несущие ген люциферазы светляка (см. рис. 9-29), не светятся. Как можно это объяснить?
8. Использование RFLР-анализа для теста на отцовство.
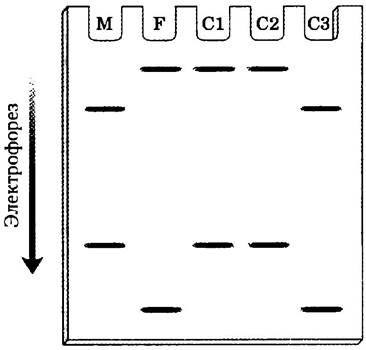
Получение отпечатков ДНК и RFLР- анализ очень часто используют при тесте па отцовство. Ребенок наследует хромосомы как от матери, так и от отца, поэтому в ДНК ребенка обнаруживаются отдельные рестрикционные фрагменты генетического материала, взятые у каждого из родителей. На основе данных электрофореза, приведенных здесь, скажите, какой ребенок, если таковой имеется, не является биологическим потомком предполагаемого отца? Обоснуйте свой вывод. Дорожка М соответствует пробе, взятой у матери, F — у предполагаемого отца, а С1, С2 и С3 — у детей.
9. Картирование хромосомного сегмента.
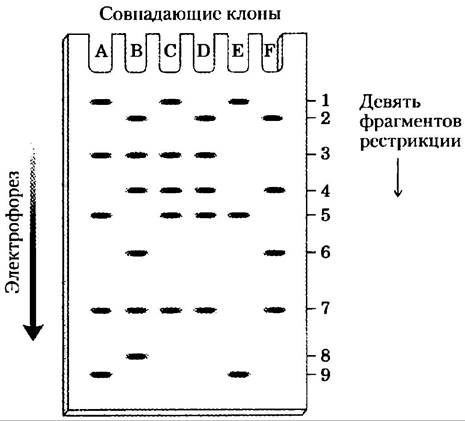
Группа перекрывающихся клонов, обозначенных буквами от А до F получена из одного и того же участка хромосомы. Каждый из клонов по отдельности разрезается рестриктазой, а фрагменты разделяются электрофорезом в агарозном геле, как это показано на рисунке снизу. Из этого участка хромосомы рестрикцией получено девять различных фрагментов, причем в каждом клоне есть их определенное подмножество. На основе этой информации определите порядок следования рестрикционных фрагментов в хромосоме.
10. Клонирование в растениях.
В методике, схематично изображенной на рис. 9-28, используются клетки бактерии р. Agrobacterium, содержащие две отдельные плазмиды. Подумайте, почему последовательности двух плазмид не объединяются в одну.
11. Получение отпечатков ДНК и RFLP- анализ.
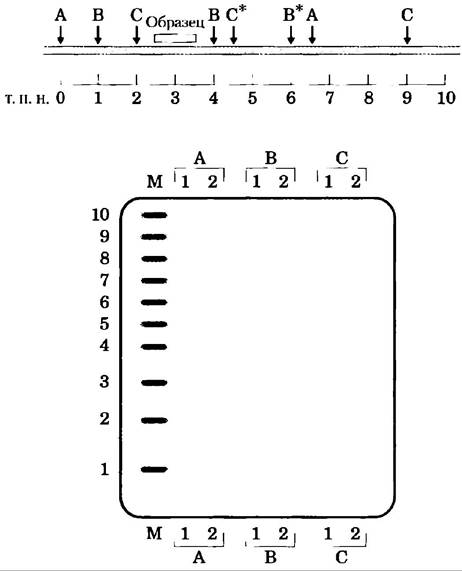
Из клеток крови двух различных людей (1 и 2) берут ДНК. В отдельных экспериментах ДНК каждого из них разрезают эндонуклеазами рестрикции А, В и С, а фрагменты разделяют электрофорезом. На рисунке изображена гипотетическая карта сегмента хромосомы человека длиной 10 000 п. н. У 2-го человека есть точечные мутации, устраняющие рестрикционные сайты распознавания В* и С*. Вы метите гель радиоактивным олигонуклеотидом, комплементарным указанной последовательности, и подносите к гелю рентгеновскую пленку. Укажите, где бы на пленке вы ожидали увидеть полосы. Дорожки в геле отмечены на сопутствующей диаграмме.
12. Использование фотолитографии в приготовлении ДНК-микрочипов.
На рис. 9-21 показаны начальные этапы в процессе приготовления ДНК-микрочипа или ДНК-чипа с использованием фотолитографии. Опишите остальные этапы, необходимые для получения требуемых последовательностей (разная четырехнуклеотидная последовательность в каждой из четырех ячеек), изображенных на рисунке на первой иллюстрации в рамке. После каждого шага покажите получившуюся нуклеотидную последовательность, прикрепленную к каждой ячейке.
13. Клонирование в млекопитающих.
Ретровирусные векторы, приведенные на рис. 9-32, позволяют осуществить эффективную интеграцию привнесенной ДНК в геном млекопитающего. Объясните, как эти векторы, у которых нет генов репликации и упаковки вируса (gag, pol, env), помещаются в инфицирующие вирусные частицы. Подумайте, почему так важно, чтобы у этих векторов не было генов репликации и упаковки.
Анализ экспериментальных данных
14. Hindi: Первая эндонуклеаза рестрикции.
Об открытии эндонуклеазы рестрикции, которую можно было использовать на практике, впервые сообщалось в двух статьях, опубликованных в 1970 г. В первой работе Смит и Уилкокс описали выделение фермента, который расщеплял двунитевую ДНК. Нуклеазную активность фермента измеряли по снижению вязкости раствора ДНК при его обработке ферментом.
а) Почему вязкость раствора ДНК уменьшается после обработки ферментом?
Для того чтобы понять, является ли фермент эндо- или экзонуклеазой, им обрабатывали ДНК, меченную 32Р, а затем добавляли трихлоруксусную кислоту (ТХУ). В таких условиях единичные нуклеотиды должны были оставаться в растворе, а олигонуклеотиды должны были выпадать в осадок.
б) При обработке меченой ДНК ферментом не образовывалось фрагментов, растворимых в ТХУ. На основании этого результата установите, являлся ли фермент эндо- или экзонуклеазой. Объясните свои рассуждения.
При расщеплении полинуклеотидной цени фосфатная группа обычно не отщепляется, а остается связанной с 5'- или 3'-концом образующегося фрагмента ДНК. Смит и Уилкокс предложили следующий протокол для определения положения фосфатной группы.
1. Обработка немеченной ДНК-нуклеазой.
2. Добавление к аликвоте продукта, полученного на стадии 1, меченного радиоактивной меткой АТР (y-[32Р] АТР), и полинуклеотид- киназы (фермент, который присоединяет y-фосфат АТР к 5'-гидроксильной группе, но не к 5'-фосфатной, 3'-гидроксильной или 3'-фосфатной группе). Измерение количества 32Р, включенного в ДНК (образец А).
3. Добавление к продукту, полученному на стадии 1, щелочной фосфатазы (фермент, удаляющий фосфатные группы с 5'- и 3'-концов), а затем полинуклеотидкиназы и y-[32Р] АТР. Измерение количества 32Р, включенного в ДНК (образец В).
в) Оказалось, что образец А обладал радиоактивностью 136 имп./мин, а образец В — 3740 ими./мин. Зная это, ответьте, оставалась ли фосфатная группа на 5'- или на 3'-конце фрагмента ДНК после обработки ферментом? Объясните свой ответ.
г) При обработке ДНК бактериофага Т7 нуклеазой появлялось около 40 фрагментов ДНК различной длины. Каким образом этот факт свидетельствует в пользу того, что фермент рестрикции узнает специфические участки ДНК, а не вносит в двойную спираль случайные разрывы?
На данном этапе исследования существовало две гипотезы: 1) фермент вносит разрывы внутри узнаваемого им участка; 2) фермент узнает определенную последовательность, но расщепляет ДНК где-то на соседнем участке. Чтобы разрешить эту проблему, Келли и Смит определяли 5'-концевые последовательности фрагментов ДНК, образующихся при расщеплении нуклеазой. Для этого был предложен следующий протокол.
1. Обработка ДНК фага Т7 ферментом рестрикции.
2. Обработка образующихся фрагментов щелочной фосфатазой для удаления 5'-фосфатных групп.
3. Обработка полученных на стадии 2 фрагментов полинуклеотидкиназой в присутствии y-[32Р]АТР для введения метки в 5'-концы фрагментов ДНК.
4. Обработка меченых фрагментов ДНКазами с целью превращения фрагментов ДНК в смесь моно-, ди- и тринуклеотидов.
5. Определение последовательности меченых моно-, ди- и тринуклеотидов путем их сравнения с олигонуклеотидами с известной последовательностью с помощью метода тонкослойной хроматографии.
Среди меченых продуктов были обнаружены следующие: мононуклеотиды А и G, динуклеотиды 5'-рАрА-3' и 5'-рGрА-3', тринуклеотиды 5' рАрАрС-3' и 5'-рGрАрС-3'.
д) Какая гипотеза о способе расщепления последовательности ДНК нуклеазой согласуется с полученными результатами? Объясните свои рассуждения.
Келли и Смит занялись определением 3'- концевых последовательностей фрагментов, образующихся при расщеплении ДНК нуклеазой. Они обнаружили смесь 5'-рТрС-3' и 5'-рТрТ-3'. Им не удалось определить последовательности ни одного тринуклеотида на 3'-конце фрагментов.
е) На основании этих данных определите последовательность, узнаваемую нуклеазой, и укажите точное место расщепления нити ДНК внутри этого фрагмента. В качестве модели для своего ответа используйте табл. 9-2.