ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 2. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ - 2014
ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
16. ЦИКЛ ЛИМОННОЙ КИСЛОТЫ
16.4. Глиоксилатный цикл
Позвоночные не могут превращать жирные кислоты или образующийся из них ацетат в углеводы. Превращения фосфоенолпирувата в пируват (с. 79) и пирувата в ацетил-СоА (рис. 16-2) характеризуются настолько значительным отрицательным изменением стандартной свободной энергии, что они практически необратимы. Если клетка не может превращать ацетат в фосфоенолпируват, то ацетат не может использоваться в качестве исходного соединения для глюконеогенеза, где из фосфоенолпирувата образуется глюкоза (см. рис. 15-11). Не обладая подобной способностью, клетка или организм не может использовать топливные молекулы или метаболиты для получения ацетата (жирные кислоты и некоторые аминокислоты) и углеводов.
Как мы отмечали при обсуждении анаплеротических реакций (табл. 16-2), фосфоенолпируват может быть синтезирован из оксалоацетата по обратимой реакции, катализируемой ФЕП- карбоксикиназой:
Оксалоацетат + GTP ⇄ фосфоенолпируват + СО2 + GDP
Поскольку атомы углерода ацетатной группы, вступающей в цикл лимонной кислоты, спустя восемь стадий оказываются в составе молекулы оксалоацетата, этот путь, казалось бы, пригоден для получения оксалоацетата из ацетата и, таким образом, может поставлять фосфоенолпируват для глюконеогенеза. Однако, как следует из стехиометрии, в цикле лимонной кислоты не происходит полного превращения ацетата в оксалоацетат; у позвоночных на каждые два углерода, вступающих в цикл в виде ацетил- СоА, два углерода покидают его в виде СО2. У многих других организмов для превращения ацетата в углеводы служит глиоксилатный цикл.
В глиоксилатном цикле из ацетата образуются четырехуглеродные соединения
Для растений, некоторых беспозвоночных и некоторых микроорганизмов (в том числе Е. coli и дрожжей) ацетат может служить и топливом, и исходным соединением для образования фосфо- енолпирувата в синтезе углеводов. У таких организмов ферменты глиоксилатного цикла катализируют полное превращение ацетата в сукцинат или другой четырехуглеродный интермедиат цикла лимонной кислоты:
2 Ацетил-СоА + NAD+ + 2Н2О —> сукцинат + 2СоА + NADH + Н+
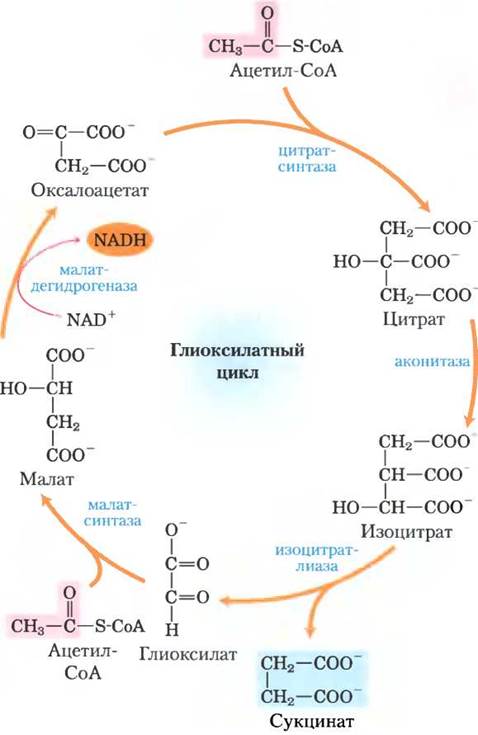
В глиоксилатном цикле ацетил-СоА вступает в реакцию конденсации с оксалоацетатом с образованием цитрата, а цитрат превращается в изоцитрат — в точности как в цикле лимонной кислоты. Однако на следующей стадии изоцитрат разлагается не под действием изоцитратдегидрогеназы, а с помощью изоцитратлиазы, в результате чего образуются сукцинат и глиоксилат. Далее глиоксилат взаимодействует со следующей молекулой ацетил-СоА с образованием малата под действием малатсинтазы. Затем малат окисляется до оксалоацетата, который может реагировать с новой молекулой ацетил-СоА, начиная новый оборот цикла (рис. 16-20). В каждом обороте глиоксилатного цикла расходуются две молекулы ацетил-СоА и образуется одна молекула сукцината, доступная для дальнейших реакций биосинтеза. Сукцинат через фумарат и малат может превращаться в оксалоацетат, который, в свою очередь, может превращаться в фосфоенолпируват под действием ФЕП-карбоксикиназы и далее в глюкозу по пути глюконеогенеза. У позвоночных нет ферментов, способных осуществлять глиоксилатный цикл (нет изоцитратлиазы и малатсинтазы), поэтому они не могут проводить синтез глюкозы из липидов.
Рис. 16-20. Глиоксилатный цикл. Цитратсинтаза, аконитаза и малатдегидрогеназа глиоксилатного цикла — изоформы соответствующих ферментов цикла лимонной кислоты. Изоцитратлиаза и малатсинтаза специфичны только для глиоксилатного цикла. Обратите внимание, что две ацетильные группы (показаны розовым цветом) входят в цикл, а четыре атома углерода в форме сукцината (выделен голубым) его покидают. Глиоксилатный цикл был исследован Хансом Корнбергом и Нейлом Мэдсеном в лаборатории Ханса Кребса.
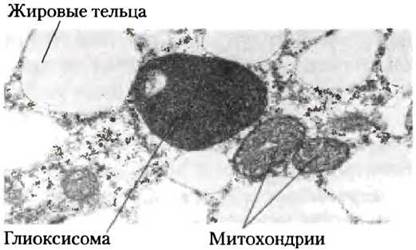
У растений ферменты глиоксилатного цикла заключены в мембраносвязанные органеллы, называемые глиоксисомами, которые представляют собой специализированные пероксисомы (рис. 16-21). Общие для цикла лимонной кислоты и глиоксилатного цикла ферменты имеют две изоформы: одна специфична для митохондрий, другая — для глиоксисом. В растениях глиоксисомы существуют не всегда. Они появляются в богатых липидами семенах в процессе прорастания, прежде чем растение приобретет способность синтезировать глюкозу путем фотосинтеза. Кроме ферментов глиоксилатного цикла в глиоксисомах содержатся все ферменты, необходимые для расщепления жирных кислот, запасенных в семенах (см. рис. 17-13). Образующийся при расщеплении липидов ацетил-СоА далее превращается в сукцинат через глиоксилатный цикл, а сукцинат транспортируется в митохондрии, где преобразуется в малат под действием ферментов цикла лимонной кислоты. Цитозольная изоформа малатдегидрогеназы окисляет малат до оксалоацетата — исходного субстрата для глюконеогенеза. Таким образом, прорастающие семена способны конвертировать запасные жиры в глюкозу.
Рис. 16-21. Электронная микрофотография прорастающего семени огурца: видны глиоксисома, митохондрии и жировые тельца.
Цикл лимонной кислоты и глиоксилатный цикл регулируются согласованно
В прорастающих семенах растений ферментативные превращения ди- и трикарбоновых кислот происходят в трех внутриклеточных отделах: в митохондриях, в глиоксисомах и в цитозоле. Между этими отделами клетки постоянно происходит обмен метаболитами (рис. 16-22).
Рис. 16-22. Связь между глиоксилатным циклом и циклом лимонной кислоты. Реакции глиоксилатного цикла (в глиоксисомах) происходят одновременно с реакциями цикла лимонной кислоты (в митохондриях) и связаны с ними; интермедиаты обоих процессов перемещаются между этими отделами клетки. Ферменты цикла лимонной кислоты катализируют превращение сукцината в оксалоацетат. Окисление жирных кислот до ацетил-СоА описано в гл. 17; синтез гексоз из оксалоацетата обсуждается в гл. 20.
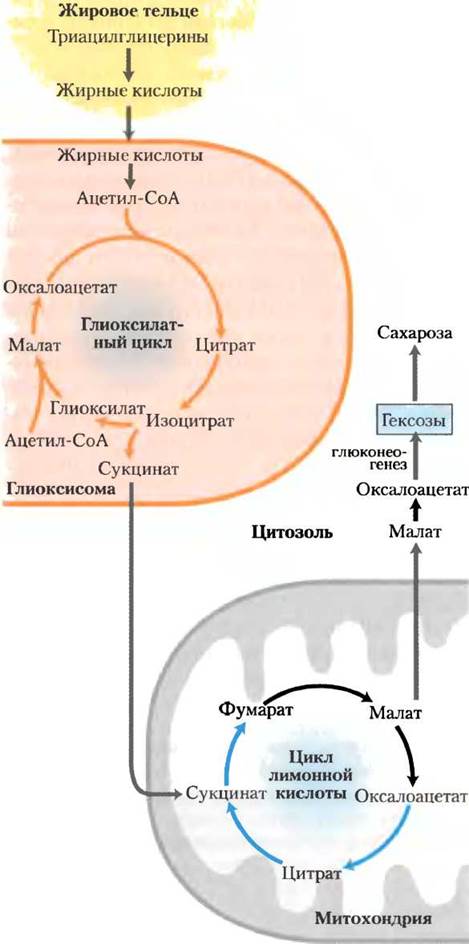
Углеродный скелет оксалоацетата из цикла лимонной кислоты (в митохондриях) переносится в глиоксисомы в форме аспартата. Аспартат превращается в оксалоацетат, который вступает в реакцию конденсации с ацетил-СоА, возникшим в результате разложения жирных кислот. Образовавшийся таким образом цитрат далее под действием аконитазы превращается в изоцитрат, а затем под действием изоцитратлиазы расщепляется на глиоксилат и сукцинат. Сукцинат возвращается в митохондрии, где вновь включается в цикл лимонной кислоты и превращается в малат, который переходит в цитозоль и под действием цитозольной формы малатдегидрогена- зы окисляется до оксалоацетата. Оксалоацетат путем глюконеогенеза преобразуется в гексозы и сахарозу, которые могут транспортироваться растущими корнями и побегами растения. В перечисленных превращениях задействованы четыре пути метаболизма: расщепление жирных кислот до ацетил-СоА (в глиоксисомах), глиоксилатный цикл (в глиоксисомах), цикл лимонной кислоты (в митохондриях) и глюконеогенез (в цитозоле).
Для использования общих интермедиатов необходимо, чтобы эти пути метаболизма регулировались согласованно. Ключевым интермедиатом в участке разветвления глиоксилатного цикла и цикла лимонной кислоты является изоцитрат (рис. 16-23). Регуляция изоцитратдегидрогеназы осуществляется путем ковалентной модификации: специфическая протеинкиназа фосфорилирует и тем самым инактивирует этот фермент. В результате этой инактивации изоцитрат направляется в глиоксилатный цикл, где он служит отправной точкой для синтеза глюкозы. Фосфопротеинфосфатаза удаляет фосфорильную группу из изоцитратдегидрогеназы, вновь активируя фермент и способствуя тому, что большее количество изоцитрата направляется в цикл лимонной кислоты для получения энергии. Активностью протеинкиназы и фосфопротеинфосфатазы обладает один и тот же полипептид.
Рис. 16-23. Согласованная регуляция глиоксилатного цикла и цикла лимонной кислоты. Регуляция активности изоцитратдегидрогеназы определяет распределение изоцитрата между двумя циклами. При инактивации фермента фосфорилированием специфической протеин- киназой изоцитрат направляется на реакции биосинтеза через глиоксилатный цикл. При активации фермента специфической фосфатазой изоцитрат поступает в цикл лимонной кислоты для образования АТР.
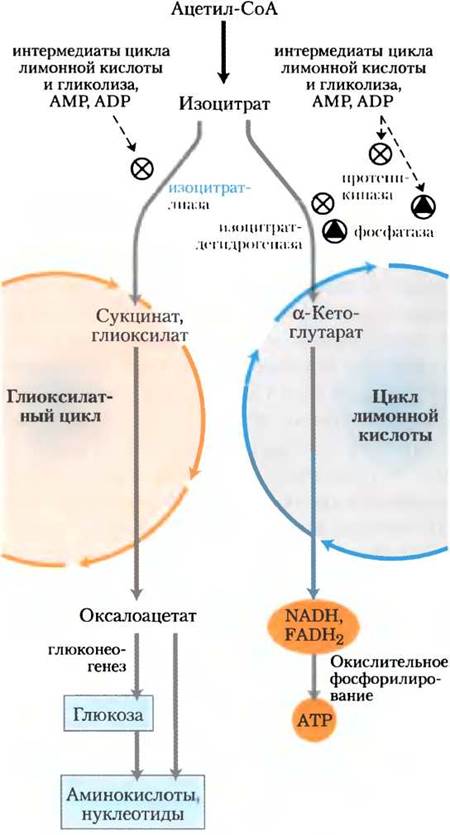
Некоторые бактерии, в том числе Е. coli, содержат полный набор ферментов глиоксилатного цикла и цикла лимонной кислоты в цитозоле и поэтому могут использовать для своего роста ацетат в качестве единственного источника углерода и энергии. В свою очередь, фосфопротеинфосфатаза, активирующая изоцитратдегидрогеназу, стимулируется интермедиатами цикла лимонной кислоты и гликолиза, а также метаболитами, накапливающимися при снижении энергетических запасов клетки (рис. 16-23). Те же метаболиты ингибируют активность протеинкиназы — бифункционального полипептида. Таким образом, появление интермедиатов центральных путей образования энергии (что сигнализирует об исчерпании клеткой энергетических запасов) приводит к активации изоцитратдегидрогеназы. При
снижении концентрации этих регуляторных молекул (означающем достаточную интенсивность потока метаболитов в цикле лимонной кислоты) изоцитратдегидрогеназа инактивируется под действием протеинкиназы.
Те же интермедиаты гликолиза и цикла лимонной кислоты, что активируют изоцитратдеги- дрогеназу, являются аллостерическими ингибиторами изоцитратлиазы. Если метаболизм, ведущий к накоплению энергии, достаточно интенсивен для поддержания на низком уровне концентраций общих интермедиатов гликолиза и цикла лимонной кислоты, изоцитратдегидрогеназа инактивируется, ингибирование изоцитратлиазы прекращается, а изоцитрат направляется в глиоксилатный цикл для дальнейшего использования в биосинтезе углеводов, аминокислот и других компонентов клетки.
Краткое содержание раздела 16.4 Глиоксилатный цикл
■ Глиоксилатный цикл реализуется в проростках семян некоторых растений и у микроорганизмов, способных использовать ацетат в качестве единственного источника углерода. В глиоксилатном цикле задействованы некоторые ферменты цикла лимонной кислоты, а также два других фермента — изоцитратлиаза и малатсинтаза.
■ Обходные реакции глиоксилатного цикла, заменяющие две стадии декарбоксилирования в цикле лимонной кислоты, делают возможным образование сукцината, оксалоацетата и других интермедиатов полностью из ацетил- СоА. Образующийся при этом оксалоацетат может быть использован для синтеза глюкозы через глюконеогенез.
■ Распределение изоцитрата между циклом лимонной кислоты и глиоксилатным циклом контролируется на уровне изоцитратдегидрогеназы, активность которой регулируется обратимым фосфорилированием.
■ У позвоночных глиоксилатный цикл не реализуется, поэтому они не могут синтезировать глюкозу из ацетата или жирных кислот через ацетил-СоА.