ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 2. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ - 2014
ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
Если вас посещает какая-то идея, ее не следует отвергать только потому, что она не укладывается в рамки господствующей теории.
Клод Бернар, An Introduction of the Study of Experimental Medicine (Введение в экспериментальную медицину), 1813
Наиболее важной и замечательной особенностью принятия хемиосмотической гипотезы, с моей точки зрения, является альтруизм и великодушие, с которым бывшие оппоненты этой гипотезы не только согласились с ней, но и единогласно возвели ее в ранг теории.
Питер Митчелл, Нобелевская речь, 1978
19. ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ И ФОТОФОСФОРИЛИРОВАНИЕ
Окислительное фосфорилирование — кульминация энергозапасающего метаболизма у аэробных организмов. Все процессы окисления при деградации углеводов, жирных кислот и аминокислот сводятся к финальной стадии клеточного дыхания, где высвобождающаяся при окислении энергия расходуется на синтез АТР. Фотофосфорилирование — процесс, в котором фотосинтезирующие организмы улавливают солнечную энергию, главный источник энергии для биосферы Земли, и направляют ее на синтез АТР. Большинство организмов синтезируют необходимый им АТР в основном именно в процессах окислительного фосфорилирования и фотофосфорилирования.
В эукариотических клетках окислительное фосфорилирование протекает в митохондриях, фотофосфорилирование — в хлоропластах. В процессе окислительного фосфорилирования как на свету, так и в темноте происходит восстановление O2 до Н2O за счет переноса электронов от коферментов NADH и FADH2. При фотофосфорилировании окисление Н2O до O2 осуществляется с участием конечного акцептора электронов NADP+ только под действием энергии света. Несмотря на различия, эти два высокоэффективных процесса преобразования энергии протекают по весьма сходным молекулярным механизмам.
Современные представления о механизме синтеза АТР в митохондриях и хлоропластах основаны на гипотезах, выдвинутых Питером Митчеллом в 1961 г. Митчелл предположил, что разница в концентрациях протонов по обе стороны мембраны, создаваемая реакциями окисления, играет роль резервуара энергии. Эта хемиосмотическая теория была принята в качестве основополагающей в биологии XX в. Хемиосмотическая теория позволяет объяснить процессы окислительного фосфорилирования и фотофосфорилирования и такие разные пути преобразования энергии, как активный перенос через мембрану и движение бактериальных жгутиков.
Окислительное фосфорилирование и фотофосфорилирование по механизму близки в трех аспектах. Во-первых, оба процесса связаны с переносом электронов по цепи переносчиков, находящихся в мембране. Во-вторых, процесс высвобождения свободной энергии при переносе электронов «вниз», т. е. экзергонический процесс, сопряжен с транспортом протонов «вверх» через непроницаемую для протонов мембрану, что приводит к накоплению свободной энергии электронов в виде трансмембранного электрохимического потенциала (с. 551, т. 1). В-третьих, поток протонов из внутримитохондриального пространства через особые белковые каналы по градиенту концентрации предоставляет свободную энергию для синтеза АТР. Его катализирует мембранный белковый комплекс АТР-синтаза, сопрягающая поток протонов с фосфорилированием ADP
Мы начнем эту главу с окислительного фосфорилирования. Прежде всего рассмотрим цепь переносчиков электронов, их организацию в большие функциональные комплексы, встроенные в структуру внутренней мембраны митохондрий, а также уделим внимание путям переноса электронов по этой цепи и сопровождающему этот перенос транспорту протонов. Затем мы остановимся на работе замечательного ферментного комплекса в ферментативной реакции, протекающий по механизму «вращательного катализа», в результате этой реакции происходит запасание энергии протонов в форме АТР; далее рассмотрены регуляторные механизмы, координирующие окислительное фосфорилирование с различными катаболическими процессами окисления топливных молекул.
Вслед за окислительным фосфорилированием в митохондриях обсудим некоторые аспекты фотофосфорилирования и прежде всего поглощение света фотосинтезирующими пигментами, механизмы переноса электронов от Н2O к NADP+ под действием света и молекулярные основы сопряжения процессов переноса электронов и протонов. Мы покажем также сходство механизмов синтеза АТР в хлоропластах и митохондриях, эволюционные основы сохранения и воспроизведения этих механизмов.
ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ
19.1. Реакции переноса электронов в митохондриях
В1948 г. Кеннеди и Ленинджер впервые показали, что процессы окислительного фосфорилирования в эукариотических клетках происходят в митохондриях. С этого важного открытия начался современный период изучения процессов биологического преобразования энергии. Митохондрии, подобно грамотрицательным бактериям, имеют две мембраны (рис. 19-1). Внешняя мембрана митохондрии сравнительно хорошо проницаема для низкомолекулярных соединений (Мr <5000) и ионов, которые свободно проходят через мембрану по каналам, образованным интегральными белками поринами. Внутренняя мембрана непроницаема для большинства небольших молекул и ионов, включая протоны Н+, и только некоторые вещества с помощью специальных переносчиков могут пройти через мембрану. Внутренняя мембрана содержит компоненты дыхательной цепи и АТР-синтазу.
Альберт Л. Ленинджер, 1917-1986

Рис. 19-1. Строение митохондрий. Внутренняя мембрана уложена в складки, или кристы, которые значительно увеличивают площадь ее поверхности. Например, на внутренней мембране митохондрий клеток печени располагается 10 000 систем переноса электронов и молекул АТР-синтазы, а на внутренней мембране митохондрий клеток сердца за счет большего числа крист — в три раза больше. Внутримитохондриальный пул коферментов и промежуточных продуктов биохимических реакций отделен от цитозольного пула физически и функционально. Митохондрии клеток беспозвоночных, растений и простейших имеют идентичную структуру, но сильно различаются по форме, размерам и степени складчатости внутренних мембран.
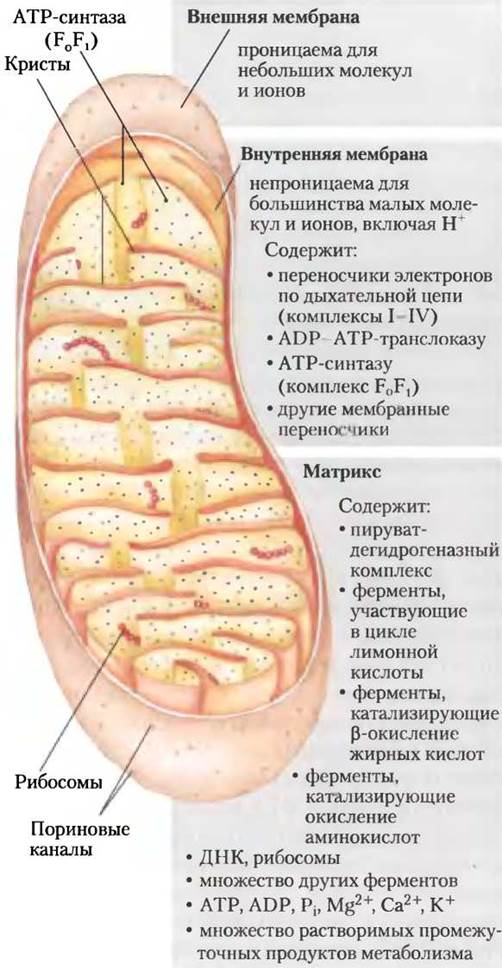
Митохондриальный матрикс, окруженный внутренней мембраной, содержит пируватдегидрогеназный комплекс и ферменты, участвующие в цикле лимонной кислоты, процессах β-окисления жирных кислот и окисления аминокислот, т. е. во всех путях окисления «топливных» молекул за исключением гликолиза, протекающего в цитозоле. Благодаря избирательной проницаемости внутренняя мембрана отделяет промежуточные соединения и ферменты, участвующие в метаболических превращениях в цитозоле, от метаболических процессов в матриксе. Внутренняя мембрана содержит и специальные переносчики, осуществляющие транспорт пирувата, жирных кислот и аминокислот или их а-кетопроизводных в матрикс, где они вовлекаются в цикл лимонной кислоты. В частности, специфические переносчики транспортируют в матрикс ADP и фосфат по мере того, как синтезированный АТР выводится из матрикса.
Электроны переносят универсальные акцепторы электронов
Окислительное фосфорилирование начинается с поступления электронов в дыхательную цепь. Большинство этих электронов возникает в результате действия дегидрогеназ, «собирающих» электроны на различных катаболических путях и доставляющих их к универсальным акцепторам электронов — никотинамиднуклеотидам (NAD+ или NADP+) или флавиннуклеотидам (FMN или FAD).
Дегидрогеназы, содержащие в качестве кофермента никотинамиднуклеотид, катализируют обратимые реакции, которые в общем виде можно записать следующим образом:
Восстановленный субстрат + NAD+ ⇄ окисленный субстрат + NADH + Н+
Восстановленный субстрат + NADP+ ⇄ окисленный субстрат + NADPH + Н+
Большинство дегидрогеназ, участвующих в процессах катаболизма, содержат кофермент NAD+ в качестве акцептора электронов. Некоторые из них локализованы в цитозоле, другие — в матриксе митохондрий, а третьи имеют как цитозольные, так и митохондриальные изоферментные формы (табл. 19-1).
Таблица 19-1. Некоторые важные реакции, катализируемые NАD(Р)Н-зависимыми дегидрогеназами
Реакция* |
Локализация** |
NAD-зависимые дегидрогеназы |
|
α-Кетоглутарат + СоА + NAD+ ⇄ сукцинил-СоА + СО2 + NADH + Н+ |
М |
L-Малат + NAD+ ⇄ оксалоацетат + NADH + Н+ |
М и Ц |
Пируват + СоА + NAD+ ⇄ ацетил-СоА + СО2 + NADH + Н+ |
М |
Глицеральдегид-3-фосфат + Pi + NAD+ ⇄ 1,3-бисфосфоглицерат + NADH + Н+ |
Ц |
Лактат + NAD+ ⇄ пируват + NADH + Н+ |
Ц |
β-Гидроксиацил-СоА + NAD+ ⇄ β-кетоацил-СоА + NADH + Н+ |
М |
NADP-зависимые дегидрогеназы |
|
Глюкозо-6-фосфат + NADP+ ⇄ 6-фосфоглюконат + NADPH + Н+ |
Ц |
NAD- или NADP-зависимые дегидрогеназы |
|
L-Глутамат + Н2O + NAD(P)+⇄ α-кетоглутарат + NH+4 + NAD(P)H |
M |
Изоцитрат + NAD(P)+ ⇄ α-кетоглутарат + СO2 + NAD(P)H + Н+ |
М и Ц |
* Эти реакции и соответствующие ферменты подробно обсуждаются в гл. 14-18.
** М — митохондрии, Ц — цитозоль.
NAD-зависимые дегидрогеназы отщепляют от окисляемых субстратов два атома водорода. Один из атомов водорода в форме гидрид-иона (:Н) присоединяется к NAD+, второй в виде иона Н+ переходит в среду (см. рис. 13-24). NADH и NADPH — водорастворимые переносчики электронов и могут в ходе реакции обратимо диссоциировать из комплекса с дегидрогеназами. NADH переносит полученные в ходе катаболических реакций электроны к началу дыхательной цепи — NADH-дегидрогеназному комплексу, описанному ниже. NADPH поставляет электроны главным образом для анаболических процессов. В клетках пулы NADH и NADPH с различными окислительно-восстановительными потенциалами строго разделены. Это достигается поддержанием отношения восстановленной формы к окисленной относительно высоким для NADPH и относительно низким для NADH. Ни NADH, ни NADPH не могут пройти через внутреннюю мембрану митохондрий, а переносимые ими электроны свободно пересекают мембрану в обоих направлениях.
Флавопротеины содержат флавиннуклеотиды FMN или FAD, связанные с белком прочными иногда ковалентными связями (см. рис. 13-27). Окисленные формы флавиннуклеотидов могут принимать от субстрата либо один электрон, переходя в форму семихинона, или два электрона — с образованием FADH2 или FMNH2. Передача электронов происходит вследствие того, что потенциал восстановления флавопротеина более положителен по сравнению с потенциалом восстановления окисляемого соединения. В отличие от коферментов NAD или NADP, стандартный потенциал восстановления флавиннуклеотидов зависит от связанного с ними белка. При взаимодействии с функциональными группами белка электронные орбитали флавинового кольца искажаются, изменяя относительную стабильность окисленной и восстановленной форм. Поэтому стандартный потенциал восстановления соответствует флавопротеину, участвующему именно в этой реакции, а не изолированным FAD или FMN. Флавиннуклеотид правильнее рассматривать как часть активного центра флавопротеина, чем как субстрат или продукт реакции переноса электронов.
Поскольку при окислительно-восстановительных реакциях флавопротеины участвуют в одно- или двухэлектронных переносах, они, по- видимому, играют роль переносчиков в реакциях с отщеплением двух электронов (как при дегидрировании) и в реакциях, где присоединяется только один электрон (как при восстановлении хинона до гидрохинона), что подробно обсуждается ниже.
Электроны проходят через цепь встроенных в мембрану переносчиков
Дыхательная цепь митохондрий состоит из ряда последовательно действующих переносчиков электронов, многие из них — интегральные белки с простетическими группами, способными присоединять или отдавать один или два электрона. В процессе окислительного фосфорилирования перенос электронов может осуществляться тремя способами. Во-первых, путем прямого переноса, как, например, при восстановлении Fe3+ до Fe2+. Во-вторых, путем переноса атома водорода (Н+ + е-) и, в-третьих, путем переноса гидрид-иона (:Н-), несущего пару электронов. В окислительно-восстановительных реакциях заряд, эквивалентный одному электрону, называют восстановительным эквивалентом.
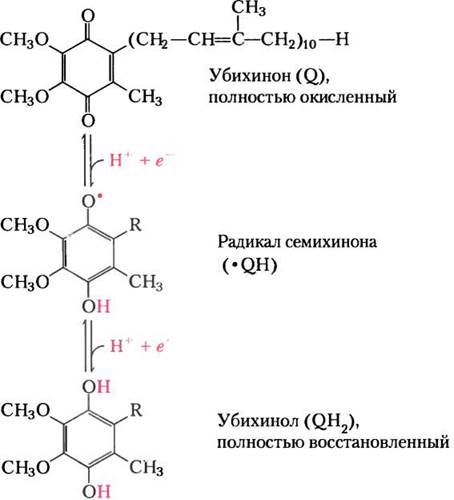
В переносе электронов в дыхательной цепи наряду с NAD и флавопротеинами участвуют гидрофобный хинон (убихинон) и железосодержащие белки двух типов — цитохромы и железосерные белки. Убихинон (кофермент Q, или просто Q) представляет собой жирорастворимый бензохинон с длинной боковой изопреноидной цепью (рис. 19-2). Аналогами митохондриального переносчика электронов кофермента Q являются пластохинон в хлоропластах растений и менахинон у бактерий. Убихинон может присоединять как один электрон, переходя в радикал семихинона (•QH), так и два электрона, восстанавливаясь до убихинола QH2(см. рис. 19-2). Подобно флавопротеинам, убихинон функционирует между донором двух электронов и акцептором одного электрона. Небольшая гидрофобная молекула убихинона способна свободно диффундировать через двойной липидный слой внутренней мембраны митохондрии и осуществлять челночный перенос восстановительных эквивалентов между другими малоподвижными переносчиками электронов, встроенными в мембрану. Убихинон переносит как электроны, так и протоны, поэтому он играет центральную роль в сопряжении переноса электронов в дыхательной цепи и транспорта протонов из матрикса митохондрий в межмембранное пространство через внутреннюю мембрану.
Рис. 19-2. Убихинон (Q, или кофермент Q). Для полного восстановления убихинона требуется два электрона и два протона. Процесс протекает в две стадии с промежуточным образованием радикала семихинона.
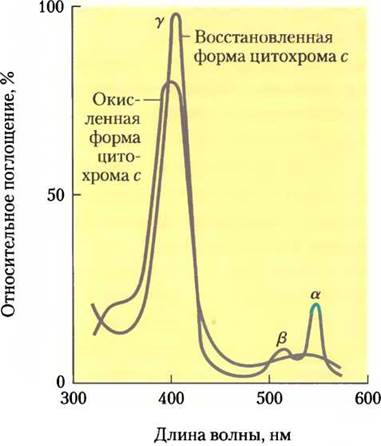
Цитохромы — это белки, имеющие характерные полосы поглощения в видимой области спектра благодаря наличию железосодержащих (гемовых) простетических групп (рис. 19-3). Митохондрии содержат цитохромы трех классов а, b и с, различающихся спектрами поглощения. Восстановленная форма всех цитохромов содержит Fe2+и имеет по три полосы поглощения в видимой области спектра (рис. 19-4). Полоса поглощения цитохрома а сдвинута в длинноволновую область — около 600 нм, цитохрома b — около 560 нм, цитохрома с — около 550 нм. Для цитохромов одного класса часто указывают в нижнем индексе длину волны максимума поглощения, например, b562.
Рис. 19-3. Простетические группы цитохромов. Простетическая группа всех цитохромов состоит из четырех пятичленных азотсодержащих колец, образующих цикл порфирина. Четыре атома азота связаны координационными связями с центральным ионом железа в восстановленном (Fе2+) или окисленном (Fе3+) состоянии. Железопротопорфирин IX найден в цитохроме b, в гемоглобине и миоглобине (см. рис. 4-16, т. 1). Гем С в цитохроме с ковалентно связан с белком через тиоэфирные связи с двумя остатками цистеина. Гем А в цитохроме а содержит длинную изопреноидную боковую цепь у одного из пятичленных колец. Характерное сильное поглощение гемов в видимой области спектра обусловлено системой сопряженных двойных связей (выделены розовым).
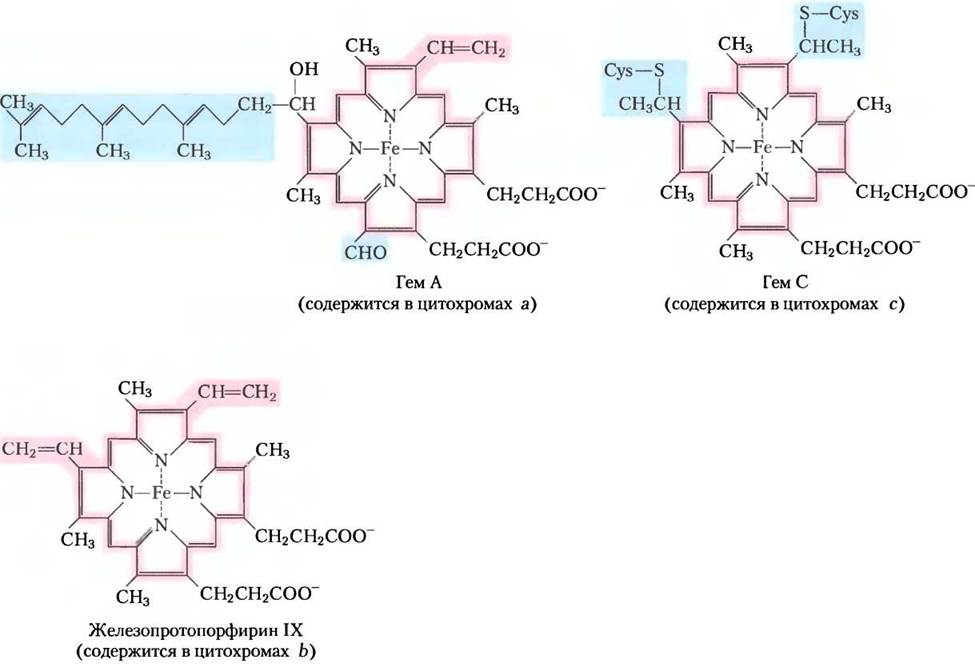
В цитохромах а и b гемовая группа связана с белком прочно, но не ковалентно. Напротив, в цитохромах с гем присоединяется к белку при помощи ковалентных связей, образованных с двумя остатками цистеина в белке (рис. 19-3). Аналогично флавопротеинам, стандартные восстановительные потенциалы гемового железа в цитохроме зависят от взаимодействия атома железа с боковыми цепями белка и вследствие этого для цитохромов различаются. Цитохромы а и b и некоторые цитохромы с представлены интегральными белками внутренней мембраны митохондрий. Исключение составляет митохондриальный цитохром с; это водорастворимый белок, удерживаемый силами электростатического притяжения на внешней стороне внутренней мембраны.
Рис. 19-4. Спектры поглощения окисленной (красная кривая) и восстановленной (синяя кривая) форм цитохрома с. α, β, y — полосы поглощения восстановленной формы.
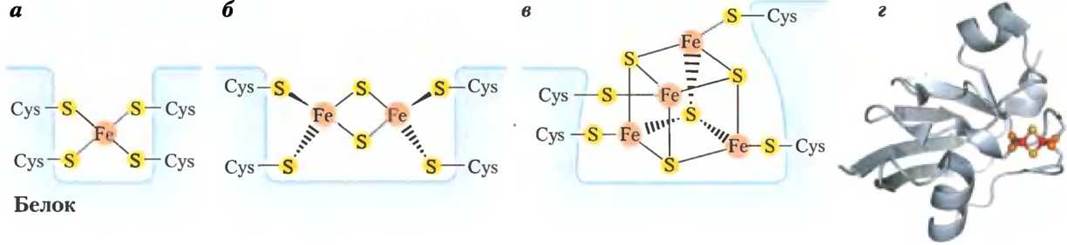
В железосерных белках атомы железа не включены в гем, а связаны либо с атомами неорганической серы, либо с атомами серы, принадлежащими остаткам цистеина в полипептидной цепи фермента, либо с атомами серы обоих типов. Строение железосерного центра может быть достаточно простым, когда там один атом Fе связан с четырьмя SН-группами цистеина, или более сложным — два или четыре атома железа (рис. 19-5). В железосерных белках Риске (открыты Джоном С. Риске) один атом железа координирует два остатка гистидина, а не цистеина. Все железосерные белки участвуют в одноэлектронных переносах, при которых в железосерном кластере окисляется или восстанавливается один атом железа. В переносе электронов в митохондриях участвуют как минимум восемь железосерных белков. Восстановительные потенциалы этих белков зависят от их микроокружения и лежат в интервале от -0,65 до +0,45 В.
Рис. 19-5. Строение железосерных центров, а — железосерный центр простого строения. Один ион железа окружен атомами серы, принадлежащими четырем остаткам цистеина в полипептидной цепи фермента, б и в — железосерные центры более сложного строения. Каждый атом железа связан как с атомами неорганической серы, так и с атомами серы, принадлежащими остаткам цистеина в белковой части фермента, г — расположение атомов железа (красные кружочки), неорганической серы (желтые кружочки) и серы в остатках цистеина (оранжевые кружочки) в железосерном центре Fе2S2 ферредоксина, выделенного из клеток цианобактерии Аnаbаеnа 7120 (РDВ 1D 1FRD). Следует отметить, что в формульных обозначениях железосерных центров указывается только число атомов неорганической серы. Например, Fе2S2 — два атома серы. На самом же деле в этом центре атом железа окружен четырьмя атомами серы — двумя атомами неорганической серы и двумя атомами серы, принадлежащими остаткам цистеина. Стандартный потенциал восстановления железа в желез-серных центрах зависит от строения центра и природы его взаимодействий с белковой частью фермента.
Перенос электронов в дыхательной цепи митохондрий можно представить следующим образом. Электроны от NАDН, сукцината или некоторых других первичных доноров попадают на флавопротеины, убихинон и далее по цепи железосерных белков и цитохромов на молекулярный кислород. Рассмотрим методы изучения переноса электронов в дыхательной цепи митохондрий и хлоропластов.
Один из методов основан на экспериментальном измерении стандартных восстановительных потенциалов переносчиков электронов, т. е. окислительно-восстановительной системы (см. табл. 19-2). Поскольку электроны самопроизвольно переходят от систем с более низким (более отрицательным) значением Е'° к системам с более высоким значением Е'°, перенос электронов от окисляемых субстратов к кисnmлороду происходит по цепочке промежуточных переносчиков в направлении увеличения восстановительного потенциала этих переносчиков. Направление потока электронов в дыхательной цепи митохондрий было установлено на основе экспериментальных значений стандартных восстановительных потенциалов для цепочки переносчиков электронов: NАDН —> Q —> цитохром b —> цитохром c1 —> цитохром с —> цитохром а —> цитохром a3 —> O2. Следует отметить, что значения стандартных восстановительных потенциалов биологических редокс-систем в условиях клетки зависят от концентрации их восстановленных и окисленных форм и не обязательно равны значениям, измеренным в стандартных условиях (см. уравнение 13-5 на с. 47).
Таблица 19-2. Стандартные восстановительные потенциалы некоторых сопряженных окислительньно-восстановительных пар, играющих важную роль в окислительном метаболизме
Окислительно-восстановительные реакции (полуреакция) |
Е'°, В |
2Н+ + 2е- —> Н2 |
-0,414 |
NАD+ + Н+ +2е- —> NАDН |
-0,320 |
NАDР+ + Н+ + 2е- —> NАDРН |
-0,324 |
NАDН-дегидрогеназа (FМN) + 2Н+ + 2е- —> NАDН-дегидрогеназа (FМNH2) |
0,30 |
Убихинон + 2Н+ + 2е- —> убихинол |
0,045 |
Цитохром b (Fе3+) + е- —> цитохром b (Fе2+) |
0.077 |
Цитохром с1 (Fе3+) + е- —> цитохром с1 (Fе2+) |
0,22 |
Цитохром с (Fе3+) + e- —> цитохром c (Fе2+) |
0,254 |
Цитохром а (Fе3+) + е- —> цитохром а (Fе2+) |
0,29 |
Цитохром а3 (Fе3+) + е- —> цитохром а3 (Fе2+) |
0,55 |
1/2 O2 + 2Н+ + 2е- —> Н2O |
0,8166 |
Второй экспериментальный метод определения цепи переносчиков электронов в дыхательной цепи митохондрий основан на укорачивании этой цепи. Для этого в отсутствие кислорода (акцептор электронов) в митохондрию вводят донор электронов. Затем в систему вводят кислород O2 и определяют скорость окисления переносчиков спектрофотометрически. Переносчик, ближайший к концу дыхательной цепи, т. е. к O2, окисляется в первую очередь, затем второй от конца цепочки переносчиков и т. д. Установленная таким образом последовательность переносчиков электронов в митохондриальной дыхательной цепи хорошо совпадает с результатами, полученными при измерении стандартных восстановительных потенциалов.
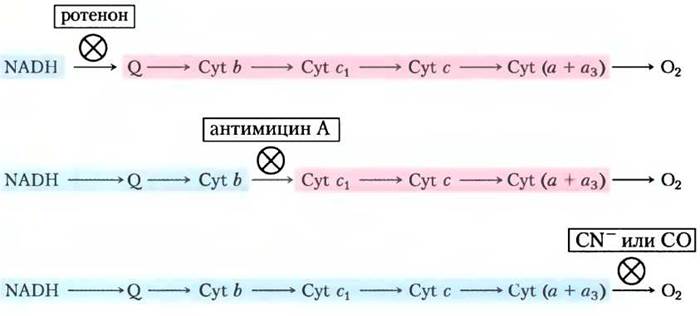
Третий метод основан на исследовании действия специфических ингибиторов, блокирующих перенос электронов на определенных участках дыхательной цепи. При введении ингибитора в цепь переноса электронов в ней возникает участок, где изменяется окислительновосстановительное состояние переносчиков. В присутствии O2 и донора электронов переносчики электронов перед заблокированным участком восстанавливаются, а сразу после этого участка — окисляются, что и регистрируют спектрофотометрически (рис. 19-6). Установленная таким образом цепь переносчиков электронов хорошо согласуется с результатами предыдущих двух методов.
Рис 19-6. Определение последовательности переносчиков электронов в дыхательной цепи митохондрий. Метод основан на изучении воздействия ингибиторов переноса электронов на степень окисления каждого переносчика. В присутствии донора электронов и кислорода O2 в качестве акцептора электронов ингибитор подавляет перенос электронов на определенных участках дыхательной цепи, изменяя окислительно-восстановительное состояние переносчиков. Переносчики, расположенные до блокированного участка, восстанавливаются (окрашены в голубой цвет), а после блокированного участка — окисляются (показаны розовым).
Переносчики электронов в дыхательной цепи митохондрий образуют полиферментные комплексы
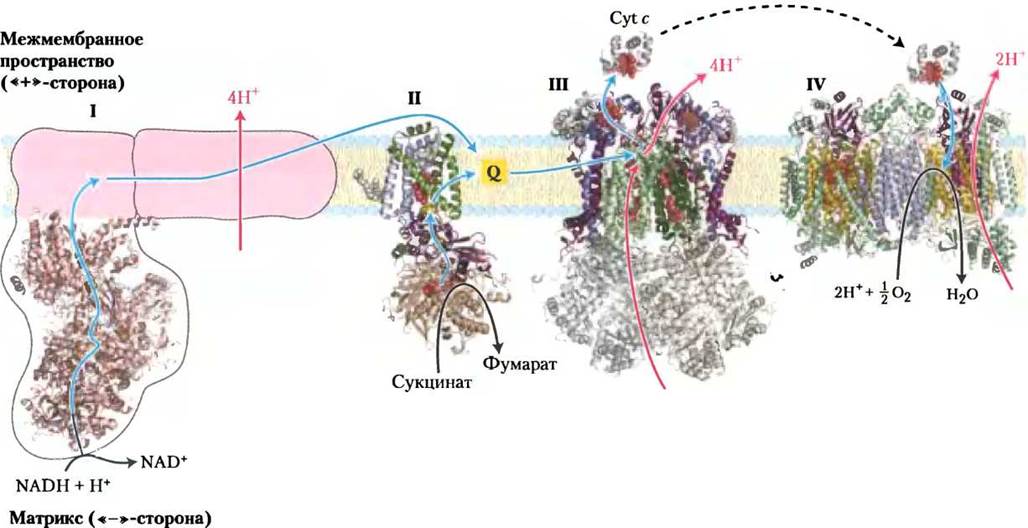
Переносчики электронов организованы в полиферментные комплексы — структурно обособленные надмолекулярные ансамбли белковых молекул, встроенные во внутреннюю мембрану. Из внутренней мембраны митохондрий путем обработки мягким детергентом удалось выделить четыре комплекса функционально связанных между собой переносчиков электронов. Каждый из них катализирует перенос электронов на определенном участке дыхательной цепи (табл. 19-3; рис. 19-7). Комплексы I и II катализируют перенос электронов на убихинон от NADH и сукцината соответственно. Комплекс III переносит электроны от восстановленной формы убихинона на цитохром с, а комплекс IV завершает передачу электронов по дыхательной цепи, передавая их от цитохрома с непосредственно на кислород.
Далее обсуждаются структурные особенности и функции белковых комплексов, участвующих в переносе электронов в дыхательной цепи митохондрий.
Таблица 19-3. Белки, участвующие в переносе электронов в дыхательной цепи митохондрий
Комплекс фермента с белком |
Масса, кДа |
Число субъединиц* |
Простетические группы |
NАDH-дегидрогеназа (комплекс I) |
850 |
43 (14) |
FMN, FeS |
Сукцинатдегидрогеназа (комплекс II) |
140 |
4 |
FAD, FeS |
Убихинон-цитохром с-оксиредуктаза (комплекс III) |
250 |
11 |
гемы, FeS |
Цитохром с** |
13 |
1 |
гем |
Цитохромоксидаза (комплекс IV) |
160 |
13 (3-4) |
гемы, CuA, CuB |
* В скобках указано число субъединиц в аналогичном комплексе, выделенном из клеток бактерий.
** Цитохром с не входит в состав ферментного комплекса; это водорастворимый мембранный белок, свободно передвигающийся между комплексами III и IV.
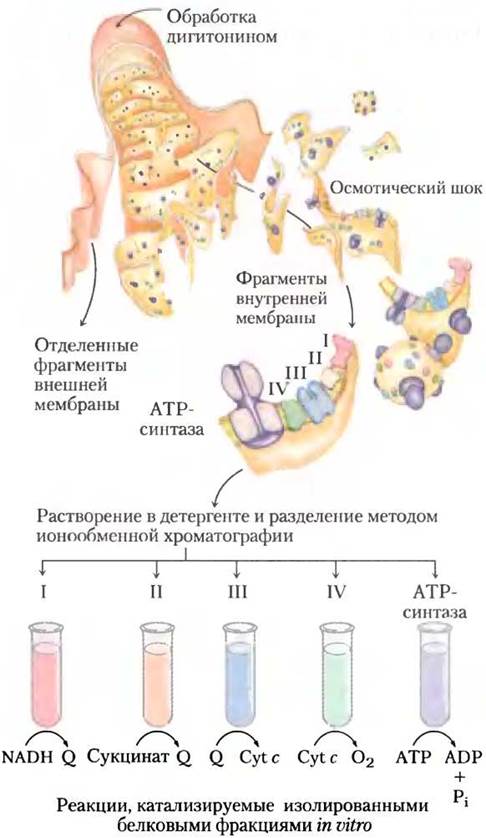
Рис. 19-7. Выделение из митохондрий ферментных комплексов, участвующих в переносе электронов в дыхательной цепи. Внешнюю мембрану митохондрий отделяют, обрабатывая митохондрии детергентом дигитонином. Внутреннюю мембрану отделяют от митохондриального матрикса методом осмотического шока. Фрагменты внутренней мембраны осторожно растворяют во втором детергенте и из этого раствора выделяют методом ионообменной хроматографии белковые комплексы I—IV, участвующие в процессах митохондриального переноса электронов, и ATP-синтазу, которую иногда называют комплексом V (см. табл. 19-3). Комплексы I—IV в изолированном состоянии катализируют передачу электронов от NADH или сукцината к промежуточным переносчикам (Q и цитохрому с), а затем к молекулярному кислороду. Изолированная АТР-синтаза обладает in vitro только ATP-гидролизующей (АТРазной) активностью, но не может катализировать синтез АТР.
Комплекс I осуществляет перенос электронов от NАDН к убихинону. Схема взаимодействия комплекса I с комплексом II и убихиноном в процессе передачи электронов представлена на рис. 19-8. Комплекс I (NАDН-убихинонредуктаза, или NADH-дeгидpoгeнaзa) состоит из 42 различных полипептидных цепей и включает в качестве простетической группы FМN и не менее шести железосерных центров. Методом электронной спектроскопии высокого разрешения установлено, что комплекс I имеет форму латинской буквы L, один конец которой погружен в матрикс, а другой прикреплен к мембране (рис. 19-9). Комплекс I катализирует два протекающих одновременно и связанных между собой процесса. Во-первых, экзергонический процесс передачи гидрид-иона от NАDН на убихинон и поглощение протона из матрикса, что можно представить в виде следующего уравнения:
NАDН + Н+ + Q —> NАD+ + QH2 (19-1)
Рис. 19-8. Перенос электронов от NАDН, сукцината, СоА-производных жирных кислот и глицерин-3- фосфата к убихинону. От NАDН электроны попадают на флавопротеин, затем на ряд железосерных белков в комплексе 1, с которого переносятся на кофермент Q. От сукцината электроны передаются через флавопротеин на железо-серные центры в комплексе II и затем на кофермент Q. Гпицерин-3- фосфат отдает электроны флавопротеину глицерин- 3-фосфатдегидрогеназе, расположенной на наружной стороне внутренней мембраны митохондрий, откуда электроны также попадают на кофермент Q. Ацил-СоА-дегидрогеназа (первый фермент в цикле β-окисления жирных кислот) отдает электроны электронпереносящему флавопротеину (ЭПФ), с которого они попадают на кофермент Q через ЭПФ- убихинон-оксиредуктазу.
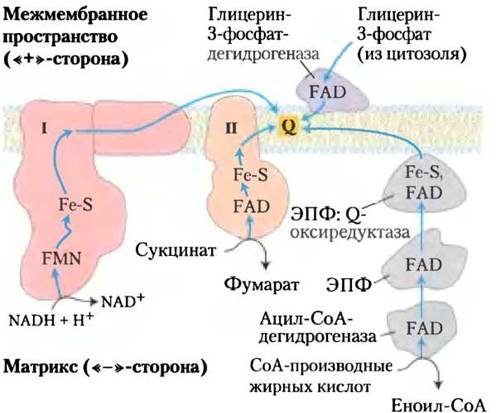
Рис. 19-9. Строение NADH-убихинон-оксидоре- дуктазы (комплекса I). Комплекс I осуществляет перенос иона водорода от NADH на FMN, от которого два электрона переносятся через ряд FeS-центров на железосерный белок N-2, локализованный в погруженной в матрикс части комплекса I. Выступающий в матрикс домен удалось кристаллизовать и изучить методом рентгеноструктурного анализа (PDB ID 2FUG); структура мембранного домена комплекса I пока неизвестна. От группы белков N-2 электроны переносятся в прикрепленную к мембране часть комплекса, где они восстанавливают убихинон до убихинола QН2, диффундирующего затем в двойной липидный слой мембраны. Перенос каждой пары электронов сопровождается выкачиванием из матрикса четырех протонов. Механизм сопряжения переноса электронов и протонов в комплексе I недостаточно ясен. Предположительно, в комплексе I, как и в комплексе III, электроны и протоны переносятся коферментом Q, совершающим особые циклические превращения, называемые Q-циклами (рис. 19-12). Молекула QН2 участвует в переносе пары электронов дважды. На поверхности внутренней мембраны в результате выкачивания протонов создается электрохимический потенциал. Внутренняя сторона внутренней мембраны заряжается отрицательно «-», внешняя — положительно «+». Некоторое количество свободной энергии, выделяющейся при переносе электронов и расходуемой на синтез АТР, запасается в виде электрохимического потенциала.
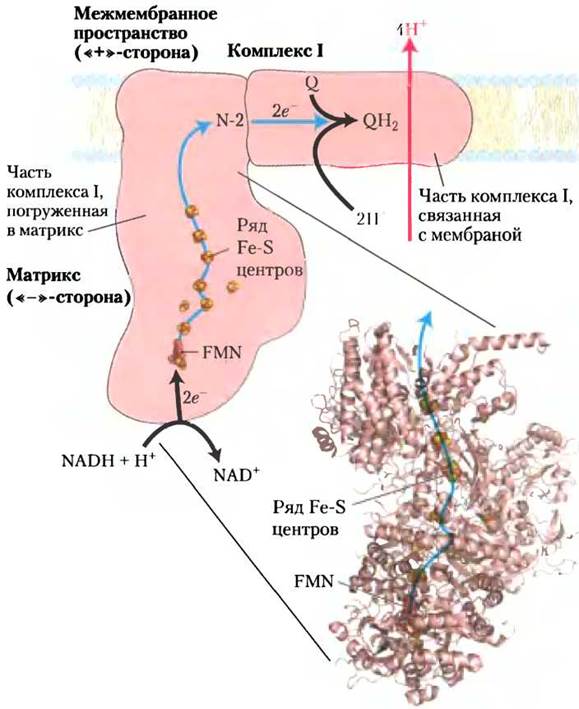
Во-вторых, эндергонический процесс выкачивания четырех протонов из матрикса и перенос их через внутреннюю мембрану в межмембранное пространство, т. е. только в одну сторону (векторно). На это затрачивается часть энергии переносимых по цепи электронов. Поскольку каждый протон несет положительный заряд, теряющий протоны матрикс заряжается отрицательно, а приобретающее протоны межмембранное пространство положительно. Чтобы подчеркнуть векторный транспорт протонов, суммарное уравнение часто записывают с соответствующими нижними индексами, указывающими местонахождение протона: «+» для положительно заряженной стороны внутренней мембраны, обращенной в межмембранное пространство, «-» для отрицательно заряженной стороны, прилегающей к матриксу.
NADH + 5Н+«-» + Q —> NAD+ + QH2 + 4H+«+» (19-2)
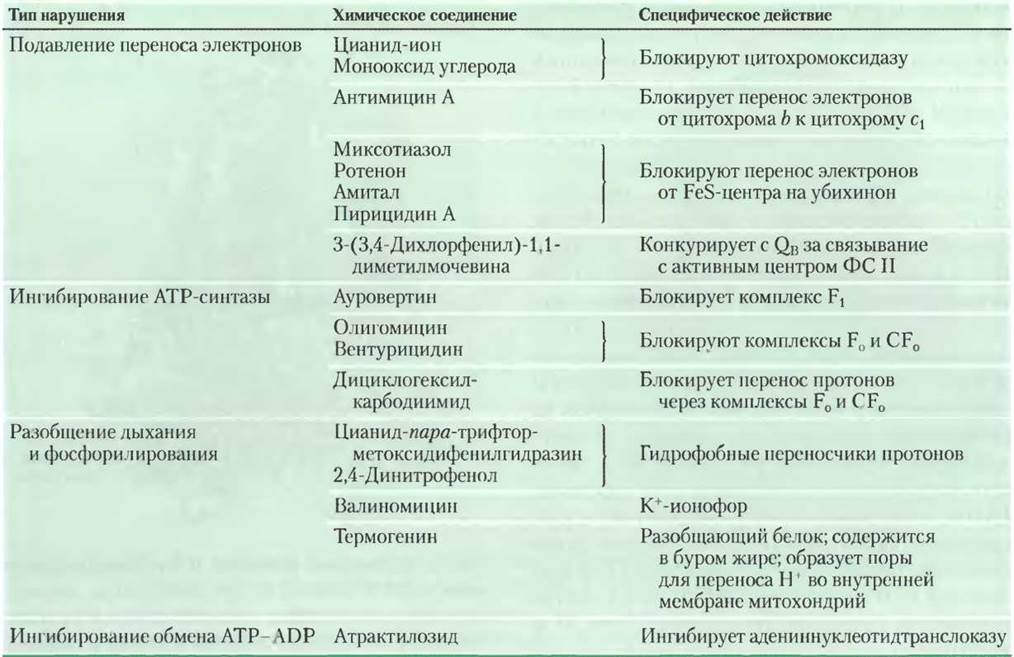
Некоторые химические соединения блокируют перенос электронов от железосерных центров комплекса I на убихинон, останавливая весь процесс окислительного фосфорилирования. Например, амитал — лекарственный препарат из группы барбитуратов, ротенон — содержащийся в растениях и используемый в качестве инсектицида, антибиотик пирицидин А (см. табл. 19-4).
Таблица 19-4. Химические соединения, нарушающие процесс окислительного фосфорилирования и фотофосфорилирования
Во внутренней митохондриальной мембране полностью восстановленная форма убихинона убихинол QH2 (рис. 19-2) диффундирует от комплекса I к комплексу III. В комплексе III при переносе протона из матрикса в межмембранное пространство QH2 окисляется до Q.
Комплекс II (сукцинатдегидрогеназа), описанный в гл. 16, переносит электроны от сукцината на убихинон Q и является единственным ферментом цикла лимонной кислоты, прочно связанным с митохондриальной мембраной (с. 198). Комплекс II имеет меньший размер, чем комплекс I, и менее сложное строение. Он включает пять простетических групп двух типов и четыре субъединицы (рис. 19-10). Субъединицы С и D комплекса II — трансмембранные интегральные белки, спирали которых трижды пересекают мембрану. В эти субъединицы включены гемовые группы типа b и центр связывания с убихиноном — конечным акцептором электронов в окислительновосстановительных реакциях, катализируемых комплексом II. Субъединицы А и В комплекса II простираются в митохондриальный матрикс (в бактериальных клетках — в цитозоль) и несут три железо-серных центра типа Fe2S2, а также прочносвязанный с апоферментом кофермент FAD и центр связывания с сукцинатом — субстратом сукцинатдегидрогеназы. Общая длина пути электронов от центра связывания с сукцинатом до FAD и затем через FeS-центры к центру связывания убихинона составляет не более 40 А. Расстояние между отдельными переносчиками электронов в этой цепи не превышает 11 А, что оптимально для быстрого переноса электронов (рис. 19-10).
Рис. 19-10. Строение комплекса II (сукцинатдегидрогеназы) (PDB ID 1NEK). Этот комплекс (показан фермент, изолированный из сердца свиньи) состоит из двух субъединиц С и одной D, пронизывающих мембрану насквозь. В цитоплазме находятся субъединицы В и А. В полипептидной цепи А после простетической группы FAD расположен центр связывания с сукцинатом. Субъединица В включает три FeS-центра. Убихинон связан с субъединицей С. Гем b расположен между субъединицами С и D. Две молекулы фосфатидил-этаноламина так прочно связаны с субъединицей D, что кристаллизуются вместе с белком. Электроны, путь которых обозначен голубыми стрелками, движутся от сукцината к FAD и далее через три железосерных центра к убихинону. Гем b не участвует в переносе электронов, но препятствует образованию активных форм кислорода (АФК) в случае «утечки» электронов.
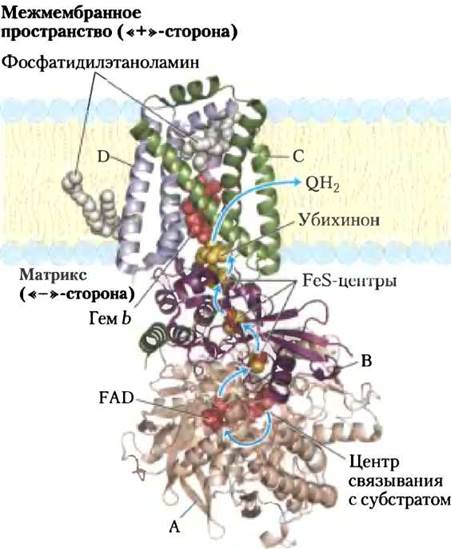
Гем b в комплексе II, по-видимому, не переносит электроны, а лишь уменьшает вероятность неполного восстановления молекулярного кислорода до Н2О, что происходит в результате неферментативного переноса («утечки») электронов на пути от сукцината к О2. «Утечка» электронов приводит к образованию токсичных для клеток активных форм кислорода (АФК): пероксида водорода и супероксидного радикала •О2-, что подробно обсуждается ниже. Причиной образования большого количества АФК в процессе окисления сукцината могут быть точечные мутации полипептидных цепей в комплексе II на участках около гема b или центра связывания с хиноном. У людей с такими мутациями развивается наследственная болезнь параганглиома — доброкачественные опухоли головы и шеи, обычно в области сонной артерии — органа, наиболее чувствительного к уровню кислорода в крови.
■ В дыхательной цепи на убихинон, минуя комплекс II, поступают электроны от всех, кроме сукцината, субстратов, окисленных митохондриальными мембранными дегидрогеназами. На первой стадии β-окисления ацил-СоА-производных жирных кислот под действием флавопротеина ацил- СоА-дегидрогеназы электроны от субстрата переносятся на FAD — простетическую группу ацил-СоА-дегидрогеназы (см. рис. 17-8). Восстановленная форма ацил-СоА-дегидрогеназы передает затем электроны специфическому переносчику электронов ЭПФ, который, в свою очередь, через ЭПФ-убихинон-оксиредуктазу переносит электроны на убихинон (рис. 19-8). Глицерин-3-фосфат, образующийся либо в ходе гликолиза при восстановлении дигидроксиацетонфосфата, либо из глицерина, выделяющегося при расщеплении триацилглицерина, окисляется глицерин-3-фосфатдегидрогеназой — ферментом из класса флавопротеинов (см. рис. 17-4). Глицерин-3-фосфатдегидрогеназа локализована на внешней стороне внутренней мембраны митохондрий и, подобно сукцинатдегидрогеназе и ацил-СоА-дегидрогеназе, передает электроны на убихинон (рис. 19-8). Роль глицерин-3-фосфат-дегидрогеназы в так называемой глицеринфосфатной челночной системе переноса восстановительных эквивалентов от цитозольного NADH в митохондриальный матрикс обсуждается в разд. 19.2 (см. рис. 19-30). Действие всех этих ферментов направлено на восстановление убихинона до убихинола QH2, который затем вновь окисляется комплесом III.
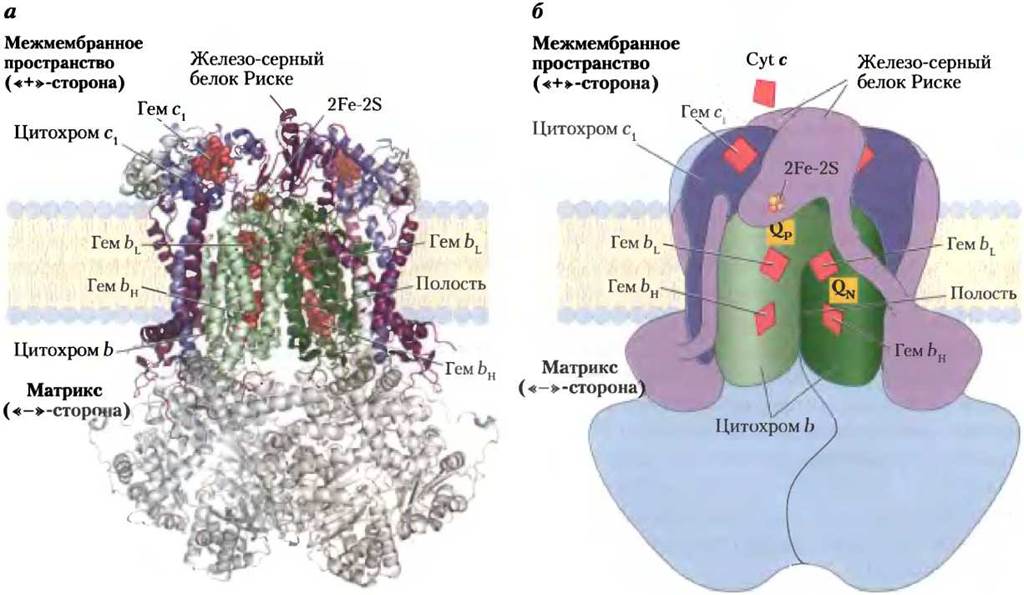
Комплекс III (комплекс цитохромов bc1, или убихинон-цитохром-с-оксиредуктаза) — следующее после комплекса II звено в дыхательной цепи; он переносит электроны от убихинона на цитохром с. Комплекс III также осуществляет связь между переносом электронов от убихинола QH2 на цитохром с и транспортом протонов из матрикса в межмембранное пространство. Полная расшифровка структуры огромного белкового комплекса III (рис. 19-11) и комплекса IV, свойства которого обсуждаются далее, была осуществлена по результатам рентгеноструктурного анализа в 1985-1998 гг. Это был важный этап в изучении процессов переноса электронов по митохондриальной дыхательной цепи и биохимических функций дыхательных комплексов.
Функциональной единицей комплекса III является димер, в котором две мономерные единицы цитохрома b окружают полость в середине мембраны, где убихинон может свободно перемещаться со стороны матрикса (участок Q«-» в одном мономере) в межмембранное пространство (участок Q«+» в другом мономере) для переноса электронов и протонов через внутреннюю мембрану митохондрий (рис. 19-11, б).
Рис. 19-11. Цитохром bс1 (комплекс III). Комплекс III включает два идентичных мономера, объединенных в функциональный ансамбль. Каждый из мономеров содержит 11 различных субъединиц, а — в функциональное ядро каждого мономера входят три субъединицы: цитохром b, (зеленая спираль), включающий гемы bH и b1, железо-серный белок Риске (фиолетовые шарики) с двумя железо-серными центрами Fe2S2и цитохром с (синяя спираль), имеющий один гем (PDB ID 1BGY). б — видно, как цитохром с1 и железо-серный белок Риске выступают над положительно заряженной поверхностью внутренней мембраны, что позволяет им взаимодействовать с цитохромом с, который расположен в межмембранном пространстве и не входит в состав комплекса III. Два центра связывания с убихиноном Q«-» и Q«+» могут присоединять два разных ингибитора, блокирующих окислительное фосфорилирование. Антибиотик антимицин А, блокирующий перенос электронов от гема bH на убихинон, присоединяется к центру Q«-», расположенному близко от гема bH на отрицательно заряженной (матриксной) стороне внутренней мембраны. Миксотиазол, блокирующий перенос электронов от убихинола QН2 на железо-серный белок Риске, присоединяется к центру Q«+», расположенному около Fe2S2-центpa и гема bLна положительно заряженной стороне внутренней мембраны. Димерная структура комплекса III важна для его каталитической активности. На границе соприкосновения двух мономеров находятся углубления, в каждом из которых расположен Q«+»-центр одного мономера и Q«-»-центр другого мономера. Промежуточные продукты окислительно-восстановительных превращений убихинона движутся внутри этих углублений.
Комплекс III может существовать в виде двух конформаций (не показано). В одной из них FeS-центр белка Риске повернут в сторону гема цитохрома c1, являющегося акцептором электронов этого белка, и удален от цитохрома b и центра связывания убихинола QН2, от которых электроны переносятся на железосерный центр белка Риске. В другой конформации FeS-центр белка Риске повернут от цитохрома с1 в сторону цитохрома b. Поскольку локализация белка Риске в комплексе III зависит от степени окисления его железосерного центра, при окислительно-восстановительных реакциях в дыхательной цепи митохондрий комплекс III переходит из одной конформации в другую.
На основе данных о строении комплекса III и результатов биохимических исследований был предложен механизм переноса электронов и протонов через комплекс III (Q-цикл).Общее уравнение окислительно-восстановительных реакций в Q-цикле (рис. 19-12) можно представить так:
QH2 + 2 Cyt c1 (окисл.) + 2H+«-» —> Q + 2 Cyt c1 (восст.) + 4H+«+» (19-3)
Рис. 19-12. Две стадии Q-цикла. Направление движения протонов через комплекс III показано голубыми стрелками. На первой стадии (слева) Q на отрицательно заряженной стороне восстанавливается, образуя радикал семихинона, который на второй стадии (справа) превращается в QН2. Две молекулы QН2 окисляются до Q на положительно заряженной стороне мембраны, высвобождая в межмембранное пространство по два протона от каждой молекулы QH2 (всего высвобождаются четыре протона). Каждая молекула QН2 передает через FеS-центр белка Риске один электрон цитохрому с1 а через цитохром b — второй протон молекуле Q, расположенной ближе к отрицательно заряженной стороне мембраны. В этих двух стадиях молекула Q восстанавливается до QН2. На восстановление одной молекулы Q расходуется два перенесенных из матрикса протона.
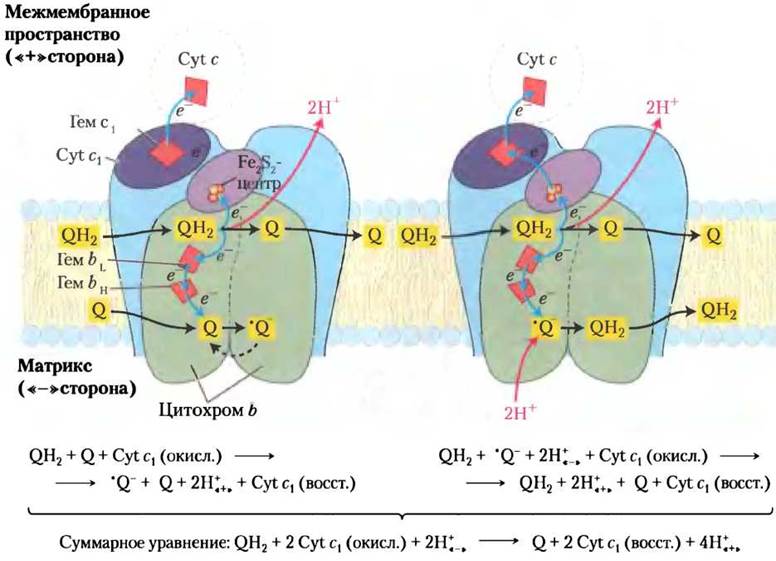
В процессе циклических превращений переносчик двух электронов убихинон (QH2) передает электроны на одноэлектронные переносчики — цитохромы b562, b566, c1 и цитохром с, а также переносит протоны из матрикса в межмембранное пространство.
При прохождении каждой пары электронов через комплекс III из матрикса выкачивается четыре протона. Сложный путь электронов через комплекс III заканчивается окислением убихинола QH2 до Q и восстановлением двух молекул цитохрома c1.
Цитохром с — водорастворимый межмембранный белок с единственной гемовой группой. После поступления электронов от комплекса III на гем цитохрома с происходит передвижение цитохрома с к комплексу IV, и цитохром с передает свои электроны на активный центр комплекса IV из двух атомов меди.
Комплекс IV (цитохромоксидаза) переносит электроны от цитохрома с на O2, восстанавливая его до Н2O и тем самым завершая перенос электронов в митохондриальной дыхательной цепи. Комплекс IV — большая белковая молекула (13 субъединиц; Мr = 204 000), локализованная во внутренней мембране митохондрий. Этот комплекс, выделенный из бактериальных клеток, состоит не менее чем из 3-4 субъединиц (при меньшем количестве нарушается его работа) (рис. 19-13).
Рис. 19-13. Строение цитохромоксидазы (комплекса IV). Этот комплекс из митохондрий клеток быка состоит из 13 субъединиц, но здесь показаны только четыре центральные субъединицы (PDB ID 1OCC). а — в комплексе IV четыре субъединицы, каждая — димер из двух одинаковых частей. Субъединица I (желтая спираль) содержит гемы а и а3 и ион меди СuB (зеленый шарик). Гем а3 и СuB образуют двухъядерный активный Fe-Cu-центр. В субъединице II (фиолетовая спираль) два иона меди соединены с белковой частью комплекса через -SH-группы остатков цистеина и образуют двухъядерный центр СuА, аналогичный Fе2S2-центрам в железосерных белках. Двухъядерный центр СиА и центр связывания с цитохромом с расположены в субъединице II на участке, выступающем в межмембранное пространство на положительно заряженной стороне внутренней мембраны. Субъединица III (сиреневая) нужна для быстрого переноса протонов через субъединицу II. Функция субъединицы IV (зеленая) пока неясна. б — двухъядерный центр СuА. Ионы меди равнозначны. При восстановлении образуется центр Cu1+Cu1+; при окислении — Cu1,5+Cu1,5+. Ионы меди окружены шестью аминокислотными остатками: два His, два Cys, Glu и Met.
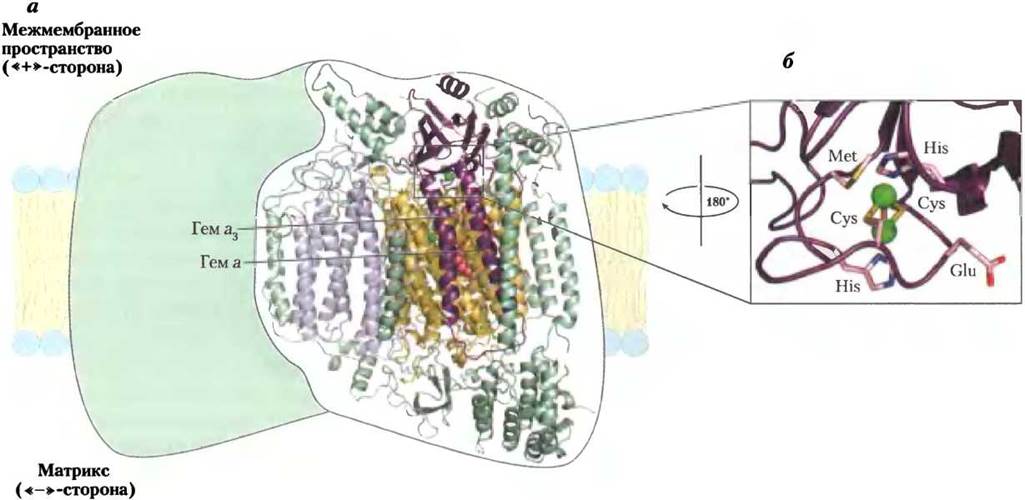
Митохондриальная субъединица II комплекса IV содержит два иона Сu, связанных через -SH-группы с двумя остатками цистеина и образующих двухъядерный центр СuА (см. рис. 19-13, б), аналогичный Fe2S2-центрам железосерных белков. Субъединица I содержит два гема (а и а3) и ион меди СuB, которые образуют второй двухъядерный центр, присоединяющий электроны от гема а и передающий их затем на молекулярный кислород, находящийся в центре связывания гема а3.
Электроны, поступившие в комплекс IV от цитохрома с, движутся от СuА-центра к гему а, затем, попав в центр а3-СuB гема, к молекулярному кислороду (рис. 19-14). Каждый раз, когда в комплекс IV поступает четыре электрона, он получает из матрикса через отрицательно заряженную поверхность мембраны также четыре протона Н+, которые отщепляются от субстратов и необходимы для превращения O2 в две молекулы Н2O. Энергия, выделяемая в этом окислительно-восстановительном процессе, используется для высвобождения протонов в межмембранное пространство (один протон на один электрон, перенесенный через комплекс IV). При этом увеличивается электрохимический потенциал, возникший в результате окислительно-восстановительных процессов и транспорта протонов в комплексах I и III. В общем виде реакции, протекающие в комплексе IV, можно записать следующим образом.
4 Cyt с (восст.) + 8Н+«-» + O2 —> 4 Cyt с (окисл.) + 4Н+«+» + 2Н2O (19-4)
Рис. 19-14. Движение электронов через комплекс IV. Субъединицы I, II и III — структурные элементы комплекса IV, обеспечивающие перенос электронов. Кроме этих полипептидов в состав комплекса IV входят 10 других (изображены зеленым цветом). Путь электронов через комплекс IV начинается с поступления электронов от каждой из двух восстановленных молекул цитохрома с (вверху) на двухъядерный центр СuА. От СuА-центра электроны передаются через гем а к Fе-Сu-центру, в который входят гем цитохрома а3 и ион СuB. Молекулярный кислород, связываясь с гемом а3, восстанавливается двумя электронами, образуя супероксид-радикал O2-2. Еще два электрона от цитохрома с (всего для образования Н2O требуется четыре электрона) превращают O2-2 в две молекулы воды. Четыре необходимых протона Н+ отщепляются от субстратов в матрикса, откуда они перекачиваются по неизвестному пока механизму.
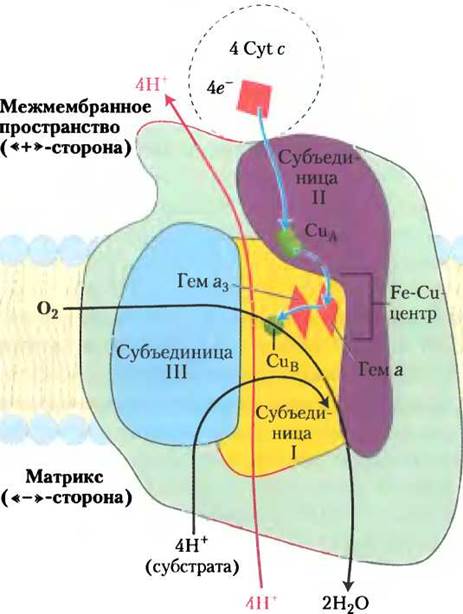
Полное восстановление молекулярного кислорода O2 происходит в окислительно-восстановительных центрах в несколько стадий, в результате одноэлектронных переносов, без высвобождения неполностью восстановленных интермедиатов, таких как пероксид водорода или гидроксильный свободный радикал, т. е. высокореакционных частиц, способных повредить клетку. Эти агрессивные частицы остаются связанными с комплексом слабыми связями до тех пор, пока полностью не завершится образование молекулы воды.
Митохондриальные комплексы могут образовывать ассоциаты — «респирасомы»
Получено много данных, подтверждающих, что в неповрежденных митохондриях комплексы дыхательной цепи прочно связаны на внутренней мембране, образуя «респирасомы» — функциональные комбинации двух или большего числа электронтранспортных комплексов. Например, при экстракции в мягких условиях мембран митохондрий комплекс III выделяется вместе с комплексом I и остается связанным с ним даже при электрофорезе в мягких условиях. Аналогично удалось выделить вместе комплексы III и IV, причем по данным электронографического исследования размеры и форма этих комплексов соответствуют распределению электронной плотности (рис. 19-15). Кинетика электронного транспорта через несколько комплексов при их тесном контакте и полном отсутствии контакта (два предельных механизма) должна быть совершенно различной. Для первого случая, когда комплексы контактируют, перенос электронов осуществляется через твердую среду. Для второго случая комплексы функционируют независимо друг от друга и электроны переносятся убихиноном и цитохромом с. Согласно полученным кинетическим данным, передача электронов происходит через твердую среду, что подтверждает модель респирасомы.
Рис. 19-15. Модель респирасомы, состоящей из комплексов III и IV. a — очищенные суперкомплексы из дрожжей, содержащие комплексы III и IV; изображение получено с помощью электронного микроскопа после окрашивания уранилацетатом. Данная электронная микрофотография получена наложением сотен электронных изображений. б — на структуру одной молекулы комплекса III (красный цвет; из дрожжей) наложена картинка электронной плотности двух молекул комплекса IV (зеленый цвет; из сердца быка), чтобы продемонстрировать возможный способ взаимодействия этих комплексов в респирасоме. Микрофотография перпендикулярно плоскости наложения структур.
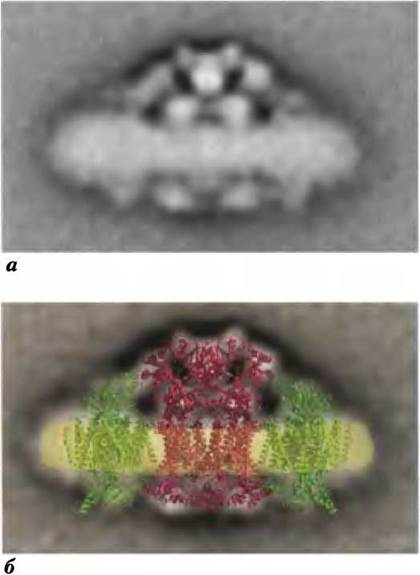
Липид кардиолипин, присутствующий в большом количестве во внутренней мембране митохондрий (см. рис. 10-9 и 11-2), возможно, играет очень важную роль в образовании респирасом. Его удаление с помощью детергентов или его отсутствие в некоторых мутантных дрожжевых клетках приводит к нарушению митохондриального транспорта электронов и потере сродства к ферментам дыхательной цепи.
Энергия, высвобождаемая при переносе электронов в дыхательной цепи, запасается в виде протонного градиента (электрохимического потенциала)
Перенос в дыхательной цепи двух электронов от восстановленного кофермента NADH на молекулярный кислород можно записать следующим образом:
NADH + Н+ + 1/2 O2 —> NAD+ + Н2O (19-5)
Этот процесс высокоэкзергонический. Стандартный восстановительный потенциал для окислительно-восстановительной пары NAD+/NADH Е'° = -0,320 В, для пары O2/Н2OЕ’° = 0,816 В. Разность стандартных потенциалов этих двух систем ∆Е'° = + 0,816 - (-0,320) = 1,14 В.
Изменение стандартной свободной энергии при переносе двух электронов от окислительновосстановительной пары NAD+/NADH к паре O2/Н2O можно рассчитать, пользуясь табл. 13-7 (с. 48):
∆G’° = -nF∆E'° (19-6)
= - 2 (96,5 кДж/В моль) • (1,14 В)
= 220 кДж/моль (NADH)
При этом предполагается, что концентрации NADH и NAD равны 1 М. Однако в активно дышащей митохондрии в результате действия многих дегидрогеназ реальное отношение концентраций [NADH] / [NAD+] оказывается значительно больше единицы, и, следовательно, изменение свободной энергии в реакции 19-5 должно быть намного больше по абсолютной величине, чем -220 кДж/моль. Аналогичные расчеты для реакции окисления сукцината показывают, что при переносе электронов от пары фумарат / сукцинат (Е’° = 0,031 В) на кислород изменение стандартной свободной энергии несколько меньше, чем в случае переноса электронов от NADH на кислород, но это также большая отрицательная величина, приблизительно равная -150 кДж/моль.
Большая часть энергии, высвобождающейся при переносе электронов от NADH на кислород, расходуется на выкачивание протонов из матрикса. При поступлении на кислород пары электронов комплекс I выкачивает из матрикса четыре протона, комплекс III — четыре Н+ и комплекс IV — два Н+ (рис. 19-16). Сопряжение процессов направленного (векторного) переноса электронов и протонов может быть описано следующим уравнением
NADH + 11 Н+«-» + 1/2 O2 —> NAD+ + 10Н+«+» + H2O (19-7)
Электрохимический потенциал, возникающий из-за разности концентраций протонов по обе стороны митохондриальной мембраны и противоположных зарядов на сторонах мембраны, является временным хранилищем большей части энергии, высвобождающейся при переносе электронов в дыхательной цепи.
Рис. 19-16. Потоки электронов и протонов через четыре комплекса в дыхательной цепи митохондрий. Электроны от NADН через комплексы I и II поступают на кофермент Q. Восстановленный убихинол QН2 — подвижный переносчик электронов и протонов. Он передает электроны комплексу III, который в свою очередь переносит их на другое подвижное звено дыхательной цепи — цитохром с. Затем комплекс IV передает электроны от восстановленного цитохрома с на молекулярный кислород O2. Перенос электронов через комплексы I, III и IV сопровождается переносом протонов из матрикса в межмембранное пространство. Через кофермент Q в дыхательную цепь поступают электроны, отщепляемые при (5-окислении жирных кислот (см. рис. 19-8). Представленные здесь структуры имеют разные источники: комплекс I — из Тhermus thermophilus (РDВ ID 2FUG); комплекс II — из сердца свиньи (РDВ ID 1ZOY); комплекс III — из сердца быка (РDВ ID) 1ВGY); цитохром с — из сердца лошади (РDВ ID 1HRG); комплекс IV — из сердца быка (РDВ ID) 1OGС).
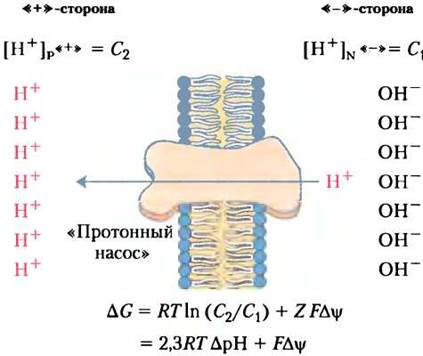
Энергия (электрохимический потенциал) мембранного градиента, так называемая протондвижущая сила, включает две составляющие. Во-первых, это химическая (потенциальная) энергия, возникшая из-за разности концентраций ионов Н+ в митохондриальном матриксе и в межмембранном пространстве. Во-вторых, электрический потенциал, обусловленный электрическим зарядом на внутренней мембране митохондрий вследствие выхода ионов Н+ из матрикса. Матрикс несет таким образом отрицательный заряд относительно межмембранного пространства (рис. 19-17).
Рис. 19-17. Возникновение протон-движущей силы. В двух компартментах митохондриального пространства, отделенных друг от друга внутренней мембраной, за счет энергии переноса электронов создается разность концентраций протонов [Н+], т. е. возникает градиент pH (∆рН) и межмембранный потенциал (∆Ψ). Результирующий эффект — протон-движущую силу (∆G) — можно рассчитать по приведенной формуле. Подробные объяснения в тексте.
Как было показано в гл. 11, изменение свободной энергии реакции переноса электронов в дыхательной цепи, когда электрохимический потенциал создается путем перехода ионов Н+ из митохондриального матрикса во внешнее пространство, можно рассчитать по следующему общему уравнению
∆G = RТIn(С2/С1) + ZF∆Ψ (19-8)
где С2 и С1 — концентрации ионов в двух компартментах, причем С2> С1, Z - заряд иона (для протона Z = 1); ∆Ψ — мембранный электрический потенциал (в вольтах).
Если принять, что переход протонов через мембрану происходит при 25 °С, то, подставляя в уравнение 19-8 соответствующие значения переменных, получаем следующее выражение
In (С2/С1) = 2,3 (Ig [Н+]«+» - Ig[Н+]«-») = 2,3 (рН«-» - pH«+») = 2,3 ∆рН
∆G = 2,3 RT∆рН + F∆Ψ = (5,70 кДж/моль) ∆рН + (96,5 кДж/В • моль) ∆Ψ (19-9)
В активно дышащей митохондрии экспериментально полученное значение мембранного потенциала составляет от 0,15 до 0,20 В, а pH матрикса примерно на 0,75 выше, чем в межмембранном пространстве.
Пример 19-1 Энергетика электронного транспорта
Рассчитайте, сколько энергии превращается в протонный градиент на внутренней митохондриальной мембране при переносе пары электронов в дыхательной цепи от NADHк кислороду. Примите, что ∆Ψ = 0,15 В, ∆рН = 0,75.
Решение. По уравнению 19-9 можно рассчитать изменение свободной энергии при переносе через внутреннюю мембрану митохондрий 1 моль протонов. Подставляя в это уравнение ∆рН = 0,75 и ∆Ψ = 0,15 В, получаем ∆G =19 кДж/моль (протонов). Согласно уравнению 19-7, перенос электронов от NADH на O2 сопровождается переходом 10 Н+. Следовательно, из 220 кДж, создаваемых при окислении 1 моль NADH, почти 200 кДж «консервируется» в виде электрохимического протонного градиента.
При спонтанном переходе протонов в матрикс «вниз» по электрохимическому градиенту выделяется свободная энергия, которая может быть использована для совершения работы. В митохондриях, хлоропластах и аэробных бактериях электрохимическая энергия протонного градиента используется для синтеза АТР из ADP и неорганического фосфата. Энергетика и стехиометрия этого процесса обсуждаются в разд. 19.2.
При окислительном фосфорилировании образуются активные формы кислорода (АФК)
На некоторых стадиях восстановления кислорода в митохондриях возможно образование реакционноспособных свободных радикалов кислорода, которые могут повреждать клетку. При передаче электронов от QH2 на комплекс III, а также от комплекса I на QH2 в качестве промежуточного продукта образуется ион-радикал •Q-. Вполне вероятно, что •Q- может передавать электрон кислороду O2
O2 + е- —> •O2-
Супероксидный ион-радикал •O2- — очень активная частица, которая может вступать в дальнейшие реакции с образованием других свободных радикалов, в частности еще более активного гидроксильного радикала •ОН (рис. 19-18).
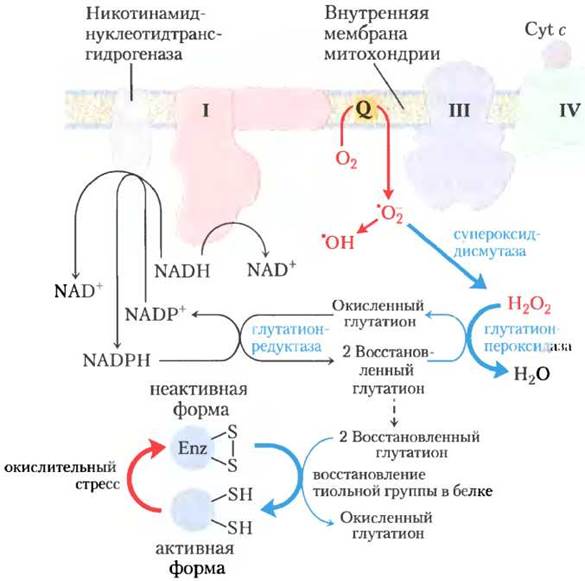
Рис. 19-18. Образование АФК в митохондриях и защита митохондрий. При нарушении скорости поступления электронов в дыхательную цепь и скорости их переноса в цепи происходит активное образование супероксидного ион-радикала •O2- в комплексах I и III, когда при неполном восстановлении радикала убихинона •Q- передает электрон молекуле O2. Супероксидный радикал действует на аконитазу, содержащую железосерный кластер 4Fе-4S, что приводит к высвобождению Fе2+. В присутствии Fе2+ происходит реакция Фентона, приводящая к образованию активного гидроксильного радикала •OН. Изображенные синим цветом реакции защищают клетки от повреждения супероксидными радикалами. Восстановленный глутатион GSН (см. рис. 22-27) дает электроны для восстановления Н2O2 и окисленных остатков Суs (-S-S-) в ферментах и других белках; восстановленная форма глутатиона регенерируется из окисленной формы GSSG в результате восстановления при участии NАDРН.
Эти активные формы кислорода могут нанести значительный вред клетке, реагируя с ферментами, мембранными липидами и нуклеиновыми кислотами, и повреждая их. В активно дышащих митохондриях от 0,1 до 4% кислорода образуют ион-радикалы •O2-, что достаточно, чтобы привести клетки к гибели, если быстро не обезвредить эти радикалы. Факторы, замедляющие поток электронов в дыхательной цепи, способствуют усилению образования супероксидного ион-радикала, возможно, из-за увеличения времени жизни •O2- в Q-цикле.
Дополнение 19-1. Горячие растения с отвратительным запахом и альтернативные пути переноса электронов
Многие цветущие растения привлекают насекомых- опылителей, распространяя пахучие вещества, которые имитируют естественный запах пищи насекомых или мест для откладывания яиц. Растения, опыляемые навозными мухами или жуками, используют для привлечения этих насекомых соединения, имитирующие запах навоза и падали.
Одно из семейств отвратительно пахнущих растений — ароидные, к ним принадлежат, например, филодендроны, аронниковые лилии и различные виды скунсовой капусты. У этих растений обычно крошечные цветки, собранные в вертикальные початки и окруженные видоизмененным листом — обверткой. Початки издают запах гниющего мяса или навоза. Кроме того, в период опыления температура початков поднимается, у некоторых видов на 20-40 °С выше температуры окружающей среды. Выделение тепла (термогенез) способствует лучшему испарению и распространению пахучих веществ и само по себе привлекает насекомых (ведь при гниении мяса или разложении навоза из-за активных метаболических процессов, вызванных гнилостными бактериями, их температура повышается). Благодаря способности к термогенезу, у восточной скунсовой капусты цветки появляются ранней весной прямо из-под снега (рис. 1).
Рис. 1. Восточная скунсовая капуста.

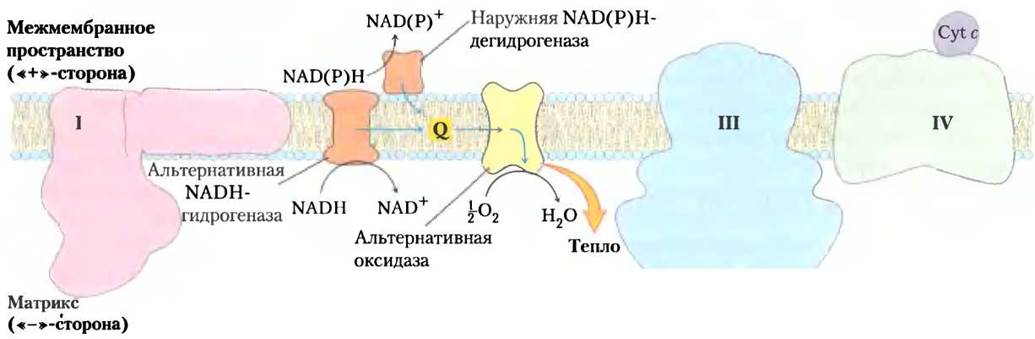
Каким же образом продуцируют тепло растения? Дело в том, что митохондрии растений, грибов и одноклеточных эукариотических организмов имеют, кроме характерных для животных цепей переноса электронов, еще альтернативную дыхательную систему, в которой энергия переноса электронов используется не для синтеза АТР, а для выработки тепла. Эта альтернативная дыхательная система имеет следующее строение. Во-первых, в ней имеется цианидрезистентная QН2-оксидаза, которая катализирует перенос электронов от убихинона прямо на молекулярный кислород, минуя комплексы III и IV (рис. 2). Во-вторых, митохондрии растений содержат ротенон-нечувствительную NADH- дегидрогеназу (см. табл. 19-4), локализованную на наружной стороне внутренней мембраны митохондрий и катализирующую перенос электронов от матриксного NADH прямо на убихинон, минуя комплекс I с его протонным насосом. В-третьих, митохондрии растений имеют еще одну NADH-дегидрогеназу на внутренней стороне внутренней митохондриальной мембраны, которая переносит электроны от цитозольного NADPH или NADH на убихинон, также минуя комплекс I. Свободная энергия, выделяемая при прохождении электронов по альтернативной дыхательной цепи, не направляется на синтез АТР, а рассеивается в виде тепла и используется растениями для выполнения различных биологических функций, например у скунсовой капусты для растапливания снега при появлении всходов и распространения зловонных запахов для привлечения насекомых-опылителей во время цветения.
Рис. 2. Пути переноса электронов во внутренней митоходриальной мембране растений. Электроны у растений могут проходить через комплексы I, II и III, как в митохондриях животных, или по альтернативной дыхательной цепи (обозначена голубыми стрелками).
Для предотвращения окислительного повреждения под действием •O2- клетка использует несколько форм фермента супероксидцисмутазы, катализирующего следующую реакцию
2 •O2- + 2 Н+ —> Н2O2 + O2
Образующийся пероксид водорода (Н2O2) обезвреживается при участии глутатионпероксидазы (рис. 19-18). Глутатионредуктаза переводит окисленный глутатион в восстановленную форму, забирая электроны от NADPH, образованного NADP-трансгидрогеназой (в митохондриях) или в пентозофосфатном пути (в цитозоле, см. рис. 14-20). Восстановленный глутатион также помогает сохранить сульфгидрильные группы белка в восстановленном состоянии, предотвращая некоторые разрушительные последствия окислительного стресса (рис. 19-18). NADP-транс- гидрогеназа играет в этом процессе очень важную роль: она приводит к образованию NADPH, необходимого для действия глутатионредуктазы.
В митохондриях растений окисление NADH протекает по другим механизмам
В периоды слабой освещенности или в темноте синтез АТР в митохондриях растений протекает по механизмам, совершенно аналогичным механизмам синтеза АТР у нефотосинтезирующих организмов. Однако на свету в митохондриях растений NADH возникает главным образом в результате превращения образовавшегося в процессе фотодыхания глицина в серин (см. рис. 20-21), что можно записать следующим образом
2 Глицина + NAD+ —> серин + СO2 + NH3 + NADH + Н+
Как показано в гл. 20, у растений накопление NADH происходит даже в том случае, если он не нужен для синтеза АТР. Из «ненужного» NADH в растениях происходит образование NAD+ в результате переноса электронов от NADH непосредственно на убихинон и затем на молекулярный кислород, минуя комплексы III и IV с их протонными «насосами». Свободная энергия переноса электронов от NADH рассеивается в виде тепла, что для некоторых растений имеет важное значение (см. доп. 19-1). Особенность переноса электронов в митохондриях растений состоит в том, что QH2-оксидаза растений, в отличие от цитохромоксидазы (комплекс IV), не ингибируется цианидами.
Краткое содержание раздела 19.1 Реакции переноса электронов в митохондриях
■ Для объяснения многих процессов, связанных с преобразованием энергии в биологических системах, включая окислительное фосфорилирование и фотофосфорилирование, предложена хемиосмотическая теория. Механизм использования энергии переноса электронов для синтеза АТР одинаков в обоих случаях. Энергия, высвобождаемая при переносе электронов, сохраняется как электрохимический градиент, возникающий в результате сопряженного с транспортом электронов «выкачивания» протонов Н+ из матрикса. Возникающая при этом протон- движущая сила используется для дальнейшего переноса протонов.
■ В митохондриях гидрид-ионы, отщепленные от субстратов NAD-зависимыми дегидрогеназами, отдают электроны в дыхательную цепь, по которой они переносятся на молекулярный кислород O2, восстанавливая его до Н2O.
■ Челночные системы доставляют восстановительные эквиваленты от цитозольного NADH к митохондриальному NADH. Во всех NAD-зависимых реакциях дегидрирования восстановительные эквиваленты переходят к митохондриальной NADH-дегидрогеназе (комплексу I).
■ Восстановительные эквиваленты переносятся через ряд железосерных центров на убихинон, который передает электроны цитохрому b — первому переносчику электронов в комплексе III. В комплексе III электроны по двум разделенным путям через два цитохрома b и цитохром c1 попадают на железосерный центр. Железосерный центр посылает по одному электрону через цитохром с на цитохромоксидазу (комплекс IV); в этом медьсодержащем ферменте присутствуют также цитохромы а и а3. Аккумулированные комплексом IV электроны передаются на молекулярный кислород O2, восстанавливая его до Н2O.
■ Электроны от различных субстратов попадают в митохондриальную цепь переносчиков электронов разными путями. Например, при окислении сукцината сукцинатдегидрогеназой (комплексом II) флавопротеин этого комплекса переносит электроны на убихинон через ряд железосерных центров. Электроны, полученные при окислении жирных кислот, переносятся на убихинон через ЭПФ.
■ Потенциально опасные активные формы кислорода (АФК), образующиеся в митохондриях, инактивируются под действием нескольких защитных ферментов, в числе которых супероксиддисмутаза и глутатион- пероксидаза.
■ У растений, грибов и одноклеточных эукариотических организмов наряду с обычным чувствительным к цианиду пути переноса электронов реализуется альтернативный нечувствительный к цианиду окислительный путь с участием NADPH.