ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 2. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ - 2014
ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
20. БИОСИНТЕЗ УГЛЕВОДОВ У РАСТЕНИЙ И БАКТЕРИЙ
20.5. Интеграция углеводного метаболизма в растительной клетке
Углеводный метаболизм в типичной растительной клетке более сложный, чем у типичной животной клетке. В растительной клетке идут те же процессы, которые производят энергию в животных клетках (гликолиз, цикл трикарбоновых кислот и окислительное фосфорилирование); в ней синтезируются гексозы из трех- или четырехуглеродных соединений путем глюконеогенеза; окисляются гексозофосфаты с образованием NАDРН (окислительный пентозофосфатный путь); и она способна создавать полимерные цепи крахмала из остатков глюкозы, соединенных α (1 —> 4)-связями, и расщеплять их до гексоз. Но помимо этих реакций с углеводами, которые присущи также животным клеткам, фотосинтезирующая растительная клетка может фиксировать СO2, включая углерод в органические соединения (ферментативная реакция под действием рубиско); использовать продукты фотосинтеза для образования триоз, гексоз и пентоз (цикл Кальвина); и превращать ацетил-СоА, образованный
при распаде жирных кислот, в четырехуглеродные соединения, а последние — в гексозы (глиоксилатный цикл и глюконеогенез соответственно). Эти уникальные процессы в растительной клетке, которые не найдены в животных клетках, разделены между несколькими компартментами: в глиоксисомах происходит глиоксилатный цикл, в хлоропластах — цикл Кальвина, в амилопластах — синтез крахмала, в вакуолях — хранение органических кислот. Интеграция событий в этих различных компартментах требует специфических транспортеров в мембране каждой органеллы, для передвижения продуктов из одной органеллы в другую или в цитозоль.
В прорастающих семенах жиры и белки превращаются в глюкозу в ходе глюконеогенеза
Многие растения запасают в семенах липиды и белки, которые необходимы при прорастании в качестве источника энергии и предшественников биосинтеза до того, как разовьется аппарат фотосинтеза. При активном глюконеогенезе в прорастающих семенах глюкоза используется для синтеза сахарозы, полисахаридов и многих метаболитов, образующихся из гексоз. В семенах растений основные количества химической энергии, необходимой для начального роста, образуется при распаде сахарозы.
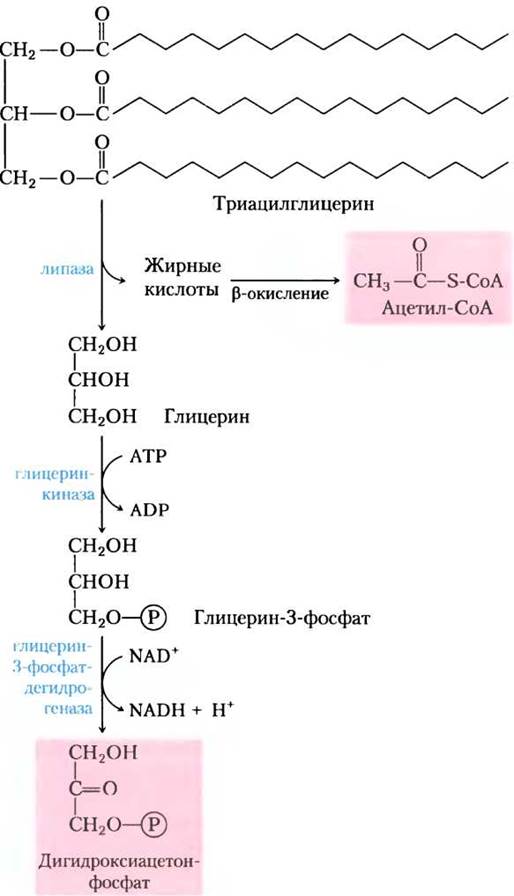
Мы отмечали ранее (гл. 14), что животные клетки могут осуществлять глюконеогенез из трех- и четырехуглеродных предшественников, но не из ацетил-СоА. Так как пируват дегидрогеназная реакция необратима (с. 182-183), клетки животных не способны превращать ацетил-СоА в пируват или оксалоацетат. В отличие от животных, растения и ряд микроорганизмов могут превращать ацетил-СоА, полученный при окислении жирных кислот, в глюкозу (рис. 20-33). Некоторые из ферментов, необходимых для этого превращения, изолированы в глиоксисомах, где специфичный для глиоксисом изозим β-окисления расщепляет жирные кислоты до ацетил-СоА (см. рис. 16-22). Физическое разделение глиоксилатного цикла и ферментов β-окисления от митохондриальных ферментов цикла трикарбоновых кислот предотвращает дальнейшее окисление ацетил-СоА до СО2. Вместо этого ацетил-СоА превращается в сукцинат в глиоксилатном цикле (см. рис. 16-20). Сукцинат проходит в матрикс митохондрий, где он под действием ферментов цикла трикарбоновых кислот превращается в оксалоацетат, который перемещается в цитозоль. В цитозоле в процессе глюконеогенеза оксалоацетат превращается в фруктозо-6-фосфат, предшественник сахарозы. Таким образом, для образования фруктозо-6-фосфата или сахарозы из запасенных липидов требуется интеграция последовательных реакций. Из-за того, что при превращениях оксалоацетата цитозоля в гексозу используются только три из четырех атомов углерода, около 75% углерода жирных кислот, запасенного в липидах семян, превращается в углеводы по комбинированным путям (рис. 20-33). Другие 25% теряются в виде СО2 при превращении оксалоацетата в фосфоенолпируват. При гидролизе запасенных триацилглицеринов образуется глицерин-3-фосфат, который может войти в путь глюконеогенеза после его окисления до дигидроксиацетонфосфата (рис. 20-34).
Рис. 20-33. Превращение запасенных жирных кислот в сахарозу в прорастающих семенах. Этот путь начинается в глиоксисомах. Образуется сукцинат, который затем экспортируется в митохондрии, где он превращается в оксалоацетат под действием ферментов цикла трикарбоновых кислот. Оксалоацетат выходит в цитозоль и служит в качестве исходного вещества для глюконеогенеза и для синтеза сахарозы — транспортной формы углерода в растениях.
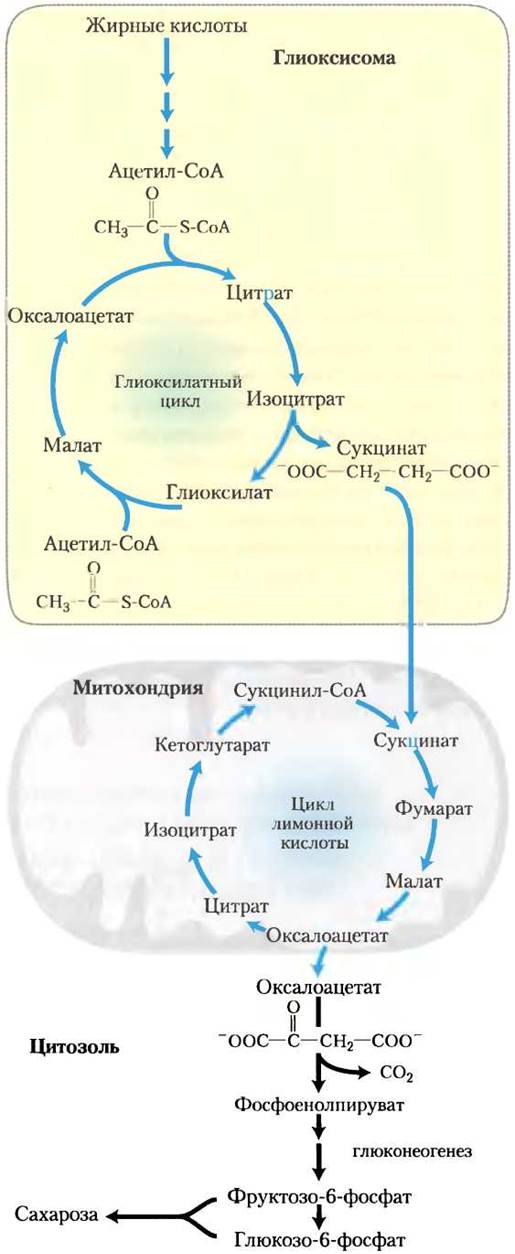
Рис. 20-34. Превращение глицериновых остатков триацилглицеринов в сахарозу в прорастающих семенах. Глицерин триацилглицеринов окисляется в дигидроксиацетонфосфат, который входит в глюконеогенез в реакции изомеризации триозофосфата.
Глюкогенные аминокислоты (см. табл. 14-4) при распаде запасных белков семян также образуют молекулы-предшественники для глюконеогенеза в реакциях трансаминирования и окисления до сукцинил-СоА, пиру вата, оксалоацетата, фумарата и α-кетоглутарата (гл. 18) — это все хороший стартовый материал для глюконеогенеза.
Метаболические пути в различных органеллах связывают пулы общих интермедиатов
Итак, мы описали отдельные реакционные цепочки метаболических превращений в растительных клетках. Однако эти реакции на самом деле взаимосвязаны, поэтому вместо последовательности реакций мы должны рассматривать пулы метаболических интермедиатов, совместно используемых метаболическими путями и связанных легкообратимыми реакциями (рис. 20-35). Один из таких пулов метаболитов включает гексозо- фосфаты: глюкозо-1-фосфат, глюкозо-6-фосфат и фруктозо-6-фосфат; второй пул — 5-фосфаты пентоз: рибозы, рибулозы и ксилулозы; третий — триозофосфаты: дигидроксиацетонфосфат и глицеральдегид-3-фосфат. Величина и направление потоков метаболитов через эти пулы меняются в ответ на изменение внешних условий, в которых находится растение, кроме того, они различны в разных типах тканей. Трансмембранные транспортеры каждой органеллы перемещают специфические соединения внутрь и наружу; регуляция этих транспортеров, возможно, влияет на обмен между пулами.
Рис. 20-35. Гексозофосфатный, пентозофосфатный и триозофосфатный пулы. Соединения в каждом пуле соединения легко взаимопревращаются, причем соответствующие реакции характеризуются очень небольшим изменением стандартной свободной энергии. Когда один из компонентов пула временно исчерпан, для его пополнения быстро устанавливается новое равновесие. Передвижение сахарофосфатов между компартментами клетки ограничено: в мембране органелл должны присутствовать специальные транспортеры.
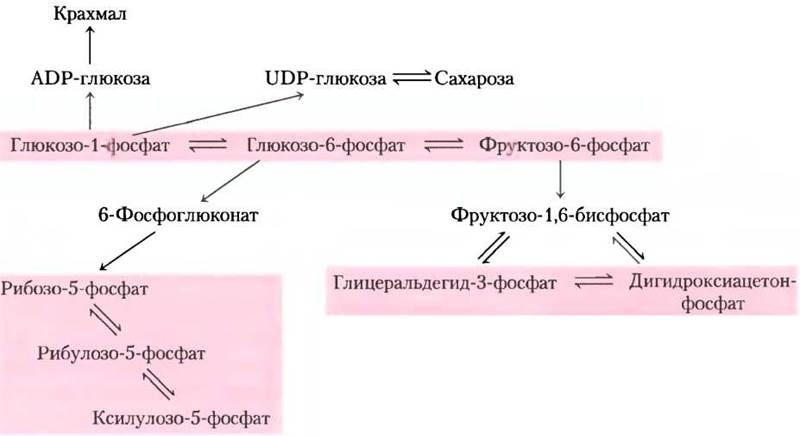
В дневное время суток триозофосфаты, образующиеся в тканях листьев в цикле Кальвина, покидают хлоропласт и попадают в гексозофосфатный пул цитозоля, где они превращаются в сахарозу для транспорта в нефотосинтезирующие ткани. В этих тканях сахароза запасается в виде крахмала и используется в качестве источника энергии в процессе гликолиза. В растущих растениях гексозофосфаты из пула расходуются и на синтез клеточной стенки. Ночью крахмал распадается и используется для получения энергии в гликолизе. Это происходит в принципе так же, как у нефотосинтезирующих организмов: образуются NАDРН и рибозо-5-фосфат по окислительному пентозофосфатному пути.
Краткое содержание раздела 20.5 Интеграция углеводного метаболизма в растительной клетке
■ Растения могут синтезировать сахара из ацетил-СоА, продукта распада жирных кислот, комбинированным действием глиоксилатного цикла и глюконеогенеза.
■ Различные пути углеводного метаболизма в растительной клетке сильно перекрываются; они используют пулы общих интермедиатов, включая гексозофосфаты, пентозофосфаты и триозофосфаты. Транспортеры в мембранах хлоропластов, митохондрий, амилопластов и пероксисом управляют передвижением сахарофосфатов между органеллами. Направления потоков метаболитов через пулы днем и ночью различны.