ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 2. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ - 2014
ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
20. БИОСИНТЕЗ УГЛЕВОДОВ У РАСТЕНИЙ И БАКТЕРИЙ
20.4. Синтез полисахаридов клеточной стенки: целлюлоза растений и пептидогликан бактерий
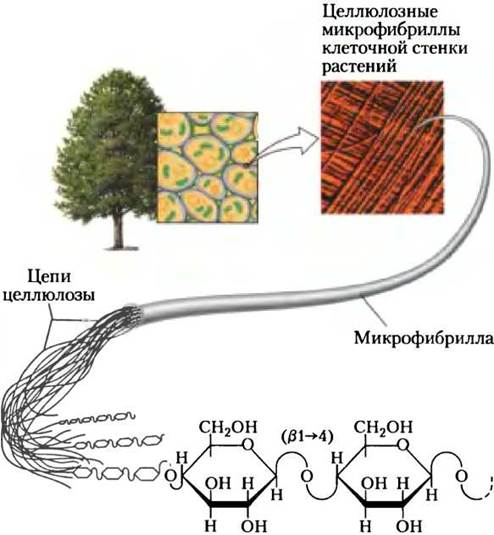
Целлюлоза — главный компонент растительной клеточной стенки, который обеспечивает ее прочность, жесткость и предохраняет клетку от набухания, а также предотвращает разрывы цитоплазматической мембраны, которые могут произойти, если под действием осмотического давления внутрь клетки войдет слишком много воды. Ежегодно на Земле растения синтезируют более 1011 т целлюлозы, что делает ее одним из самых распространенных соединений в биосфере. Целлюлоза имеет простое строение; ее линейная полимерная цепь состоит из тысяч остатков D-глюкозы, соединенных β (1 —> 4)-связями; полимерные цепи объединены в пучки из -36 параллельных друг другу нитей, которые образуют микрофибриллы (рис. 20-29).
Рис. 20-29. Строение целлюлозы. Клеточная стенка растительной клетки состоит из молекул целлюлозы, параллельные тяжи которой уложены в микрофибриллы. Большие пучки микрофибрилл, собранных из целлюлозных волокон, видны в сканирующий электронный микроскоп как нити диаметром от 5 до 12 нм, образующие на поверхности клетки несколько слоев, различимых из-за разных ориентаций волокон.
Биосинтез целлюлозы изучен в меньшей степени, чем синтез гликогена или крахмала. Как главный компонент растительной стенки целлюлоза должна синтезироваться из внутриклеточных предшественников, но откладываться и собираться на внешней стороне плазматической мембраны. Ферментативные комплексы для инициации, удлинения и экспорта цепей целлюлозы сложнее, чем для синтеза крахмала или гликогена (которые не экспортируются). Бактерии сталкиваются с похожими проблемами при синтезе комплекса полисахаридов, из которых сделана их клеточная стенка, поэтому, скорее всего, они используют те же механизмы для решения этих проблем.
Целлюлоза синтезируется надмолекулярными структурами в плазматической мембране
Сложные ферментные комплексы, которые собирают цепи целлюлозы, пронизывают плазматическую мембрану; одним концом эти комплексы связываются с субстратом UDР-глюкозой в цитозоле, а другой их конец, который отвечает за удлинение молекул целлюлозы и их кристаллизацию, выступают наружу, во внеклеточное пространство. Электронные микрофотографии, полученные методом замораживания-скалывания, показывают, что эти терминальные комплексы, также называемые розетками, состоят из шести больших частиц, образующих правильные шестиугольники (рис. 20-30). Терминальный комплекс составляют несколько белков, включая каталитическую субъединицу целлюлозосинтазы. Недавние успехи в изучении синтеза целлюлозы в основном обязаны генетическим и молекулярногенетическим исследованиям растения Аrabidорsistraliana, которое подвергли тщательному генетическому анализу, в результате геном этого растения был расшифрован. Было клонировано семейство генов, ответственных за синтез целлюлозы; выяснилось, что эти гены кодируют белки с восемью трансмембранными фрагментами и центральным доменом на цитозольной стороне плазматической мембраны, часть последовательности которого соответствует последовательности гликозилтрансферазы (рис. 20-30).
Рис. 20-30. Одна из моделей, описывающих синтез целлюлозы у сосудистых растений. Данная схема построена на основании комбинации генетических и биохимических данных, полученных для Arabidopsis thaliana и для других сосудистых растений.
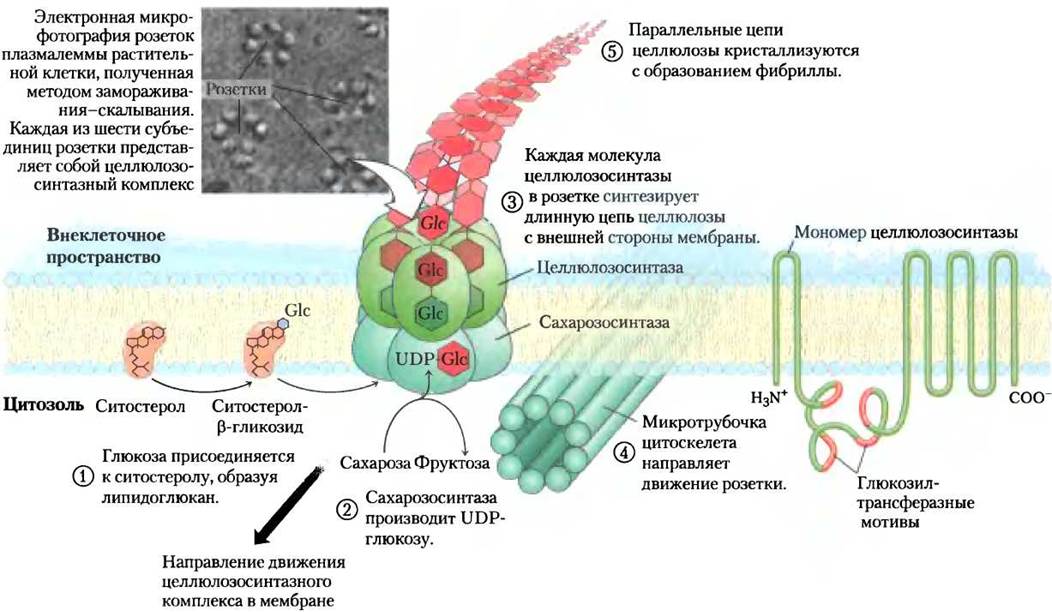
Согласно одной из принятых моделей, описывающих синтез целлюлозы, синтез новых цепей целлюлозы инициируется образованием интермедиатов, содержащих связанные липиды, что отличает синтез целлюлозы от синтеза крахмала или гликогена. Как показано на рис. 20-30 (стадия (1)), глюкоза переносится от UDP-глюкозы на мембранный липид, возможно, на растительный стерин ситостерол на внутренней стороне плазматической мембраны. Здесь внутриклеточная целлюлозосинтаза присоединяет несколько остатков глюкозы к первому остатку глюкозы β (1 —> 4)-связыо, образуя короткую олигосахаридную цепь, соединенную со ситостеролом (ситостеролдекстрином). Затем ситостеролдекстрин переворачивается, пересекает мембрану поперек и на наружной стороне клетки связывается с другой формой целлюлозосинтазы.
(2) Для синтеза целлюлозы используется UDP-глюкоза, которая образуется из сахарозы, синтезируемой при фотосинтезе в реакции, катализируемой сахарозосинтазой (названа по обратной реакции):
Сахароза + UDP —> UDP-глюкоза + фруктоза
Целлюлозосинтаза пересекает плазматическую мембрану и использует цитоплазматическую UDP-глюкозу, полученную из сахарозы в качестве предшественника для синтеза внеклеточной целлюлозы. Одна форма сахаросинтазы — мембраносвязанная сахарозосинтаза — образует комплекс с целлюлозосинтазой, поставляя UDP- глюкозу из сахарозы в синтез клеточной стенки.
(3) Другая форма целлюлозосинтазы синтезирует полимер из 500-15 000 остатков глюкозы на поверхности клетки. Работа фермента продолжается довольно долго: одна молекула фермента способна добавить большое количество остатков глюкозы, прежде чем появится растущая цепь целлюлозы. Направление роста цепи (т. е. куда присоединяется остаток глюкозы — на восстанавливающий или на невосстанавливающий конец) пока не установлено.
Каждая из шести глобул розетки состоит из многих белковых субъединиц, которые вместе синтезируют шесть цепей целлюлозы. Крупный ферментный комплекс, катализирующий этот процесс, продвигается вдоль плазматической мембраны, следуя направлению микротрубочек кортекса — цитоплазматического слоя, расположенного непосредственно под мембраной. Поскольку микротрубочки лежат в перпендикулярном направлении к оси роста растения, микрофибриллы целлюлозы укладываются перпендикулярно оси роста. Считается, что передвижение целлюлозосинтазного комплекса осуществляется за счет энергии, высвобождающейся в реакции полимеризации, а не за счет действия молекулярного мотора кинезина.
Молекулы целлюлозы собраны в кристаллические микрофибриллы (рис. 20-29), в каждой содержится 36 параллельных целлюлозных нитей с одинаковой ориентацией восстанавливающих и невосстанавливающих концов. По-видимому, каждая частица в розетке синтезирует по шесть целлюлозных цепей, поэтому на внешней поверхности клетки появляются одновременно 36 длинных полимерных молекул, которые (5) готовы к кристаллизации в микрофибриллы клеточной стенки. Когда 36 целлюлозных цепей достигают некоторой критической длины, их синтез прерывается по неизвестному механизму, затем следует кристаллизация в микрофибриллы.
В активированном предшественнике целлюлозы (в UDP-глюкозе) глюкоза образует α-связь с нуклеотидом, а в продукте (в целлюлозе) остатки глюкозы соединены β (1 —> 4)-связью, поэтому аномерный атом углерода (С-1) претерпевает инверсию конфигурации и образуется гликозидная связь. Гликозилтрансферазы, которые изменяют конфигурацию, используют механизм мономолекулярного замещения с нуклеофильной атакой на акцептор донором (UDP-глюкозой).
Некоторые бактерии (pp. Acetobacter, Agrobacteria, Rhizobia и Sarcina) и многие простые эукариоты также осуществляют синтез целлюлозы, очевидно, по похожему механизму. Но если для начала новой цепи бактерия использует мембранный липид, это не может быть стерол — бактерии не содержат стеролы.
Олигосахариды, которые связаны с липидами, — предшественники бактериальной клеточной стенки
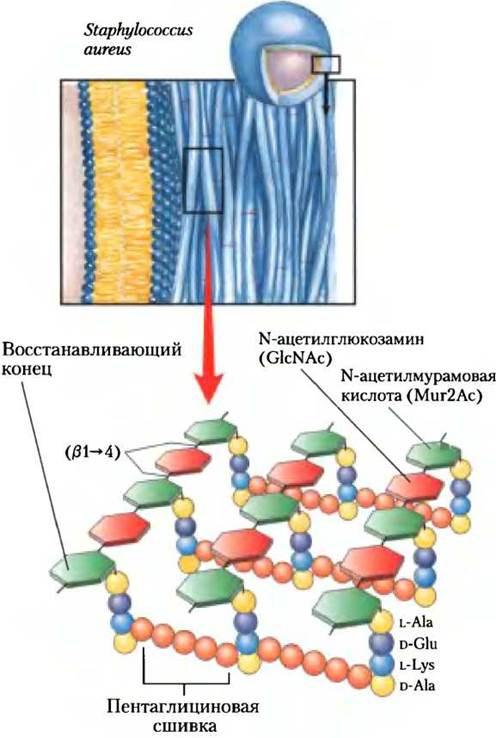
Как и растения, многие бактерии имеют прочную и толстую клеточную стенку, которая защищает их от осмотического лизиса. Пептидогликан, который придает бактериальной оболочке прочность и жесткость, представляет собой линейный сополимер из чередующихся звеньев N-ацетилглюкоза- мина (GlcNAc) и N-ацетилмурамовой кислоты (Мur2Ас), связанных β (1 —> 4)-гликозидной связью; цепи в этом сополимере (поперечно) сшиты между собой короткими пептидами, присоединенными к Мur2Ас (рис. 20-31). В процессе сборки этой сложной макромолекулы и GlcNАс, и Мur2Ас активируются путем присоединения к аномерному атому углерода уридинового нуклеотида. Сначала GlсNКАс-1-фосфат вступает в реакцию конденсации с UТР, в результате образуется UDР-GlcNАс (рис. 20-32, стадия (1)), который далее с фосфоенолпируватом дает UDР-Мur2Ас (стадия (2)); затем присоединяется пять аминокислот (стадия (D). Мur2Ас-пентапептид переносится на мембранный липид долихол (длинноцепочечный изопреноидный спирт, см. рис. 10-22, е) (стадия (4)), после чего к Мur2Ас присоединяется GlcNAc (стадия (5)). У многих бактерий к аминогруппе остатка Lys пентапептида через пептидную связь присоединяется пять молекул глицина (стадия (6)). Наконец, этот дисахаридный декапептид присоединяется к невосстанавливающему концу молекулы-праймера — пептидогликану (стадия (7)). Соседние цепи полисахаридов поперечно сшиваются в транспептидазной реакции (стадия (8)), в результате вокруг бактериальной клетки образуется прочная стенка (оболочка). Большинство эффективных антибиотиков, которые сейчас используются, ингибируют реакции синтеза пептидогликана.
Рис. 20-31. Структура пептидогликана. Показан пептидогликан клеточной стенки грамположительной бактерии Staphylococcus aureus. Пептиды (последовательность из раскрашенных сфер) ковалентно связаны с остатком N-ацетилмурамовой кислоты соседней полисахаридной цепи. Обратите внимание на то, что пептиды образованы из смеси L- и D-аминокислот. Грамположительные бактерии, такие как 5. aureus, сшиты пентаглициновы- ми цепочками. Грамотрицательные бактерии, такие как Е. coli, не содержат пентаглицинов; вместо этого терминальный остаток D-Ala тетрапептида присоединен прямо к соседнему тетрапептиду либо через L-Lys, либо через похожую на лизин диаминопимелиновую кислоту. Образование пептидной связи с остатком глутамата здесь происходит необычно — через карбоксильную группу боковой цепи.
Многие другие олигосахариды и полисахариды синтезируются по похожему механизму, в котором сахара активируются, присоединяя нуклеотиды в последовательных реакциях. Например, в гликозилировании белков (см., например, рис. 27-39, т. 3) предшественники углеводной части белковой молекулы включают нуклеотидсахара и липидсвязанные олигосахариды.
Рис. 20-32. Синтез бактериального пептидогликана. На ранних стадиях этого пути (с (1) по (4)) N-ацетилглюкозамин (GlсNAс) и N-ацетилмурамовая кислота (Мur2Ас) активируются присоединением уридинового нуклеотида (UDР) к аномерному атому углерода, а в случае Мur2Ас длинноцепочечного изопреноидного спирта (долихола), через фосфодиэфирную связь. Эти активирующие группы участвуют в образовании гликозидных связей; они служат в качестве превосходных уходящих групп. После этого происходит ((5), (6)) ассоциация дисахарида с боковой пептидной цепью (10 аминокислотных остатков), (7) этот предшественник переносится на невосстанавливающий конец существующей пептидогликановой цепочки, которая служит праймером для реакции полимеризации. Наконец, (8) в транспептидазной реакции между боковыми пептидными цепями двух соседних молекул пептидогликана остаток Glу на конце одной цепи замещается D-Аlа другой цепи, образуя поперечные сшивки. Транспептидазная реакция ингибируется пенициллинами, которые убивают бактерии, разрушая их клеточную стенку.

Краткое содержание раздела 20.4 Синтез полисахаридов клеточной стенки: целлюлоза растений и пептидогликан бактерий
■ Синтез целлюлозы происходит в терминальных комплексах (розетках) на плазматической мембране. Сначала цепи целлюлозы внутри клетки превращаются в декстрин (стерин). Затем они переворачиваются так, что олигосахаридная часть оказывается вне клетки; она переносится на целлюлозосинтазу в розетке, где потом и удлиняется. На каждой розетке одновременно образуется 36 целлюлозных цепочек с параллельной ориентацией. Цепи целлюлозы кристаллизуются в микрофибриллы, которые образуют клеточную стенку.
■ В синтезе пептидогликана бактериальной клеточной стенки также участвуют связанные с липидами олигосахариды, которые образуются внутри клетки, но потом уже в мембране макромолекула поворачивается и олигосахаридные хвосты оказываются вне клетки.