Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Биосинтез углеводов в животных тканях
Особенно активный глюконеогенез свойствен жвачным животным
Мы знаем, например, что лабораторные крысы по своему метаболизму очень близки к человеку, но это можно сказать далеко не про всех хорошо известных нам животных. У жвачных животных. в том числе у крупного рогатого скота, растительная пища, перевариваясь, подвергается ферментации в рубце под действием находящихся там бактерий. Рубцом называется первый отдел четырехкамерного желудка жвачных. У коровы объем рубца достигает 70 л. Это как бы огромный ферментёр (рис. 20-6), в котором различные обитающие здесь виды бактерий расщепляют главные компоненты растительной пищи, в первую очередь целлюлозу (гидролизовать ее неспособен ни один из обычных пищеварительных ферментов животного организма). Живущие в рубце бактерии расщепляют целлюлозу (в которой остатки глюкозы соединены ß(1→ 4)-связями) с образованием свободной D-глюкозы. На этом, однако, деятельность бактерий не кончается. Они сбраживают почти всю глюкозу до лактата и ряда других продуктов, среди которых главную роль играют ацетат, пропионат и бутират. В сутки у коровы поступает из кишечника в кровь не более нескольких граммов несброженной глюкозы. Однако корова, точно так же как крыса или человек, нуждается в глюкозе крови. Она нужна ей не только как клеточное топливо для мозга и других тканей, но также и в качестве предшественника лактозы (молочного сахара) в период лактации.
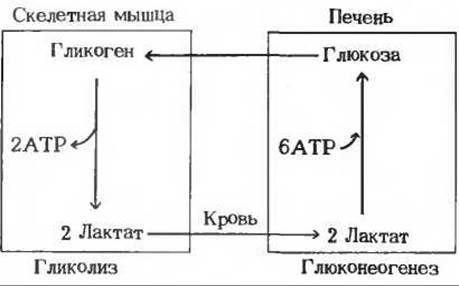
Рис. 20-5 Взаимодействие скелетных мышц и печени в процессе восстановления после тяжелой мышечной работы, во время которой происходит анаэробное расщепление гликогена с образованием двух молекул лактата и двух молекул АТР на каждую расщепленную глюкозную единицу. В период восстановления (показано красным) лактат, поступивший из мышц в кровь, превращается в печени в глюкозу крови. На образование одной молекулы глюкозы из двух молекул лактата расходуется шесть молекул АТР. Глюкоза доставляется кровью обратно в мышцы и откладывается здесь в запас в виде гликогена.
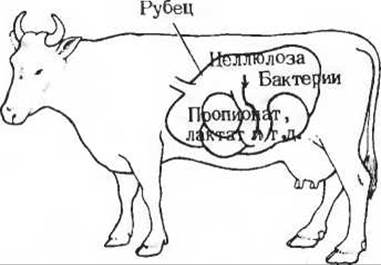
Рис. 20-6. Рубец занимающий значительную часть брюшной полости коровы, представляет собой как бы огромный ферментёр, в котором бактерии расщепляют целлюлозу ферментативным путем до глюкозы. Эта глюкоза сбраживается затем с образованием лактата, пропионата, ацетата и бутирата, которые всасываются в кровь. В печени лактат и пропионат вновь быстро превращаются в глюкозу.
Откуда же берется у коровы эта необходимая ей глюкоза, если перевариваемые углеводы почти нацело расщепляются у нее в рубце до короткоцепочечных органических кислот? Оказывается, организм коровы зависит в этом смысле от глюконеогенеза, протекающего в печени животного весьма интенсивно. Лактат, образуемый в рубце бактериями, всасывается в кровь и в печени превращается в глюкозу по описанному выше пути, точно так же как это происходит у человека или у крысы. Другой важный продукт сбраживания глюкозы в рубце, трехуглеродный пропионат (разд. 18.8), превращается в глюкозу в результате процесса, который свойствен как жвачным, так и нежвачным, но у первых играет значительно более важную (в количественном смысле) роль. Этот путь (рис. 20-7) интересен в двух отношениях: 1) в нем имеется этап, на котором двуокись углерода «фиксируется», т. е. переходит в органическую форму в результате карбоксилирования пропионил-СоА; 2) глюконеогенный путь, ведущий от пропионата к глюкозе, включает этап, катализируемый ферментом, содержащим в качестве простетической группы прочно связанную коферментную форму витамина В12-дезоксиаденозилкобаламин (разд. 10.11). Этот фермент называется метилмалонил-СоА-мутазой (разд. 18.8). В реакции, которую он катализирует, сложная замещенная алкильная группа переносится от одного атома углерода к другому, соседнему, в обмен на атом водорода, что приводит к образованию сукцинил-СоА (рис. 20-7). Возникший таким путем сукцинил-СоА превращается в малат, являющийся предшественником фосфоенолпирувата (рис. 20-2), и в конечном счете - в D-глюкозу. Следует отметить, что СО2, фиксированная при карбоксилировании пропионил-СоА, позже снова теряется. У нежвачных животных образование глюкозы из пропионата идет далеко не столь интенсивно, потому что пропионат образуется у них только в результате окисления жирных кислот с нечетным числом атомов углерода (разд. 18.8), а также в процессе окислительного расщепления трех аминокислот-метионина, изолейцина и валина.
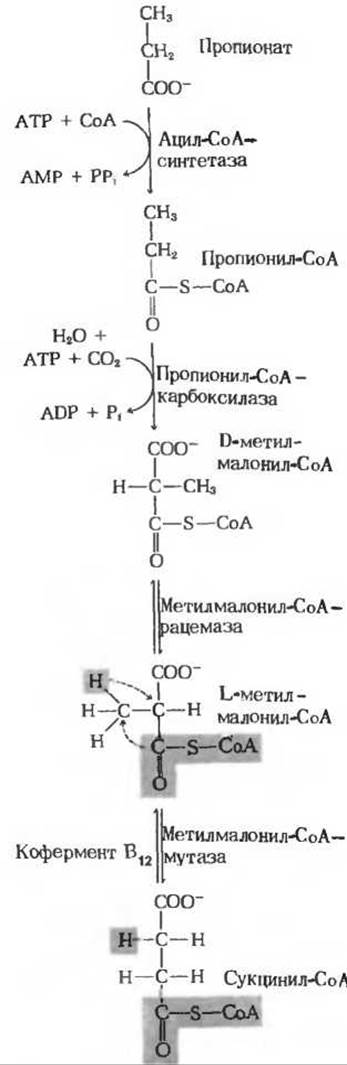
Рис. 20-7. Превращение пропионата в сукцинил-СоА, который затем может превратиться в фосфоенолпируват и в конечном счете в глюкозу. СО2, присоединившаяся при образовании D-метилмалонил-СоА, снова отщепляется, когда оксалоацетат превращается в фосфоенолпируват. Обратите внимание на обмен заместителями при двух соседних атомах углерода в молекуле L-метилмалонил-СоА. катализируемый метилмалонил-СоА—мутазой, для действия которой требуется кофермент В12. См. также разд. 18.8.